Koncept genetiky kvantitativních znaků
Fenotypová hodnota (P) vlastnosti je výsledkem působení genotypové hodnoty (G) a prostředí (E). Z grafů popisujících kvantitativní vlastnosti vyplývá hlavní koncept kvantitativní genetiky (~ příčiny způsobující variabilitu vlastnosti), který navrhl Johannsen již v roce 1909:
- fenotyp = genetické faktory + faktory prostředí
- P = G + E
Změřená fenotypová hodnota (P) je výsledkem působení genotypové hodnoty organizmu (G) a prostředí (E), ve kterém vznikla.
Genetické faktory
Aditivní působení genů
Jedná se o nejrozšířenější model dědičnosti u kvantitativních vlastností. Každá alela má specifickou metrickou hodnotu, která je přičítána ve výsledném fenotypu. Tato složka genotypu je příčinou podobnosti mezi příbuznými jedinci a je nejvíce ovlivňována fenotypovou selekcí.
Neaditivní působení genů
Dominantní genové působení - Dominantní homozygot a heterozygot přispívají stejnou mírou na fenotyp (intragenové interakce). Patří sem i neúplná dominance, superdominance a kodominance.
Genové interakce - Interakce dvou či více genů na různých lokusech kontrolujících jednu vlastnost (epistáze) se stejným nebo rozdílným účinkem (intergenové interakce). Čím více je genů pro vlastnost, tím více je možných interakcí.
Aditivní model dědičnosti
Zásadní otázkou je, jak se stává, že skutečná diskontinuální variance zapříčiněná genetickou segregací, je překládána do kontinuální variance u metrických vlastností?
Každý fenotyp vyplývá z působení různého počtu genů s aditivním působením různého počtu alel. Na obrázku níže je zobrazen případ kvalitativní vlastnosti, podmíněné různým počtem genů, bez vlivu prostředí! Čím více genů se účastní determinace vlastnosti, tím více se dosahuje kontinuálnosti (snižuje se rozdílnost diskrétních tříd).
Vliv počtu genů na fenotypovou variabilitu:
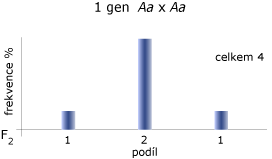
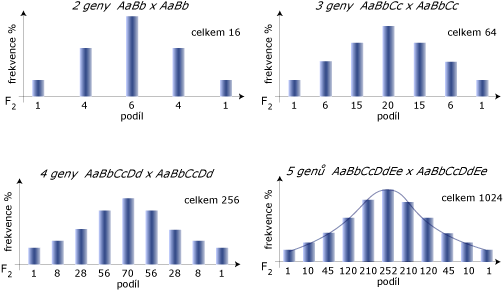
Jsou zde dva důvody:
- současná segregace mnoha genů ovlivňující vlastnost
- velká část skutečné kontinuální variance je způsobena negenetickými příčinami.
Předpokládejme, že segreguje 6 nevázaných lokusů, každý se dvěma alelami s frekvencí 0,5 a s úplnou dominancí jedné alely nad druhou na každém lokusu a že dominantní alely každého lokusu přispívají jednou jednotkou měření určité vlastnosti. Kdyby měly tvořit variabilitu jen tyto segregující geny, pak bychom získali 7 diskrétních tříd podle měření vlastnosti. To je podmíněno tím, zda jedinec nese dominantní alelu na žádném, 1., 2., ... nebo 6. lokusu. Očekávaná distribuce za současné segregace dvou alel na každém lokusu je popsána na obrázku 2 pro případ s 6 a s 24 lokusy. Každý lokus, je-li recesivně homozygotní, snižuje měření o jednu jednotku (6 lokusů) a nebo o 1⁄4 jednotky (24 lokusů). Na ose x jsou počty homozygotních lokusů s recesivní alelou a na ose y je pravděpodobnost (%) jedinců očekávané v každé třídě. Pravděpodobnosti jsou odvozeny z binomického rozložení (1⁄4 + 3⁄4)n, kde n je počet lokusů.
Kdybychom měřili opravdu přesně, tak bychom mohli odlišit jednotlivé třídy. Čím více segregujících lokusů s malým efektem, tím více tříd s menšími rozdíly mezi sebou by vzniklo. Vezmeme-li v úvahu, že na kvantitativní vlastnosti působí prostředí, tak nelze rozpoznat diskontinuitu a vytváří se kontinuální proměnlivost. Na úrovni genetické lze uvažovat o diskontinuitě, ale pozorovaná, fenotypová variabilita je kontinuální.
Očekávaná distribuce za současné segregace dvou alel na každém lokusu:
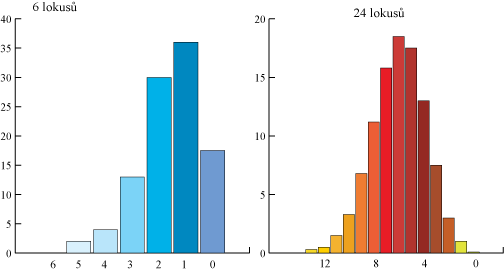
Rozlišení mezi geny s mendelistickou dědičností a geny s polygenní dědičností spočívá ve velikosti jejich efektů ve vztahu k dalším zdrojům variability. Geny s velkým účinkem (majorgeny) vytvářejí rozpoznatelnou diskontinuitu i při současné segregaci dalších genů a negenetickou varianci lze studovat metodami klasické genetiky. Geny malého účinku (minorgeny) nemohou významně přispět k diskontinuitě a nemohou být studovány individuálně. V genomu jsou i geny s intermediárním účinkem a nemohou být zahrnuty do skupiny genů s velkým nebo malým účinkem. Variance zapříčiněná současnou segregací mnoha genů se nazývá polygenní variance a geny malého účinku se označují jako polygeny.
| Počet lokusů | 1 | 2 | 3 | n |
|---|---|---|---|---|
| Počet gamet hybridů | 2 | 4 | 8 | 2n |
| Typy F1 gamet | A, a | AB, Ab, aB, ab | ABC, ABc, AbC, Abc, aBC, aBc, abC, abc | |
| Počet F2 genotypů (nebo fenotypů při aditivitě) | 3 | 9 | 27 | 3n |
| Typy F2 genotypů | AA, Aa, aa | AABB, AABb, AAbb, AaBB, AaBb, Aabb, aaBB, aaBb, aabb | AABBCC, AABBCc, AABBcc, AABbCC, AABbCc, AABbcc, AAbbCC, AAbbCc, AAbbcc, AaBBCC, AaBBCc, AaBBcc, AaBbCC, AaBbCc, AaBbcc, AabbCC, AabbCc, Aabbcc, aaBBCC, aaBBCc, aaBBcc, aaBbCC, aaBbCc, aaBbcc, aabbCC, aabbCc, aabbcc | |
| Počet různých F2 fenotypů (při úplné dominanci) | 2 | 4 | 8 | 2n |
| Počet různých F2 fenotypů (při neúplné dominanci) | 3 | 5 | 7 | 2n + 1 |
| Podíl homozygota F2 | 1/4 (AA nebo aa) | 1/16 (AABB nebo aabb) | 1/64 (AABBCC nebo aabbcc) | 1/4n |
| Fenotypový poměr F2 | 1 : 2 : 1 | 1 : 4 : 6 : 4 : 1 | 1:6:15:20:15:6:1 | (A+a)2 |
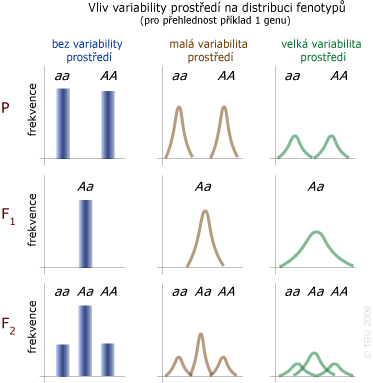
Vliv variability prostředí na celkovou variabilitu fenotypovou
Kombinováním efektů genotypové variability a variability prostředí dochází ke zvýšení variability fenotypové s kontinuálním průběhem.
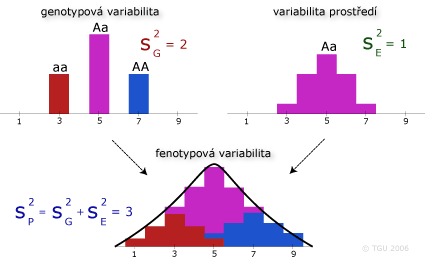
Různá úroveň variability prostředí ovlivňuje (modifikuje) rozdílně stejné genotypy.

Aktualizováno: 03.02.2015