Nenáhodné oplození
- asortativní oplození
- inbríding
Asortativní oplození
Pokud je výběr partnera závislý na určitých fenotypových vlastnostech (výška, barvy, pach…), pak se o takovém oplození říká, že je asortativní. Asortativní oplození znamená nenáhodné oplození založené na určitých fenotypových vlastnostech.
Pozitivní asortativní oplození (kromě negativního asortativního oplození) nemění frekvence alel, ale spíše mění způsob, jak jsou alely distribuovány do diploidních genotypů. Při pozitivním asortativním oplození se páří spíše fenotypově podobní jedinci, než by se dělo při náhodném oplození. Toto oplození je docela běžné. Vysocí lidé si často berou vysoké partnery a lidé menšího vzrůstu zase menší partnery. Lidé si vybírají partnery také asortativně podle různých stavů socioekonomického postavení, IQ, ... Tedy pářící se páry mají v průměru podobné fenotypy než by se dalo očekávat při náhodném spojování.
Genetickým efektem pozitivního asortativního oplození je zvýšení homozygotnosti (zvýšení frekvence homozygotních genotypů) a snížení heterozygotnosti. Např. lokus s úplnou dominancí u můry, kde můry genotypu AA mají větší délku než můry Aa, které jsou zase delší než můry genotypu aa. Pozitivní asortativní oplození se projevuje tím, že můry AA mají tendenci se pářit s můrami stejného genotypu a můry genotypu aa zase s můrami aa. Všechny tyto kombinace tvoří pouze homozygotní potomstvo. Také oplození heterozygotů mezi sebou tvoří polovinu homozygotních potomků. Přitom se vyskytuje jen málo oplození typu AA x aa, které by produkovalo heterozygotní potomstvo. Takže se frekvence heterozygotů snižuje.
Genetické efekty jsou pouze na těch lokusech, které ovlivňují fenotypovou vlastnost, na základě kterých se organizmy asortativně páří (a lokusy na ně vázané). Lokusy, které neovlivňují vlastnost, podle které si vybírají partnery, nejsou ovlivněny, protože oplození je náhodné pro genotypy na těchto lokusech. Tím se odlišuje asortativní oplození od inbrídingu, který ovlivňuje všechny lokusy stejně. Podobně jako inbríding, pozitivní asortativní oplození mění genotypové frekvence, ale bez změny frekvencí alel.
Negativní asortativní oplození (disasortativní oplození) se děje, když se páří fenotypově méně podobní jedinci mezi sebou, než by se očekávalo při náhodném oplození. Dobrým příkladem je vlastnost pohlaví - samci se páří se samicemi a naopak. Tento typ oplození je méně běžný než pozitivní asortativní oplození a to hlavně u zvířat. Dobře prostudované negativní asortativní oplození u rostlin je self-incompability (sebenesnášenlivost). Mnoho rostlinných druhů mají specifické self-incompatibility lokusy. Pylová zrna, která mají společné s rostlinami alely na takovýchto lokusech, nemohou úspěšně oplodnit ostatní rostliny se stejnými lokusy. Tím je zajištěno, že nemůže dojít k samoopylení nebo k opylení mezi úzce příbuznými rostlinami, neboť ty mají mnoho společných alel.
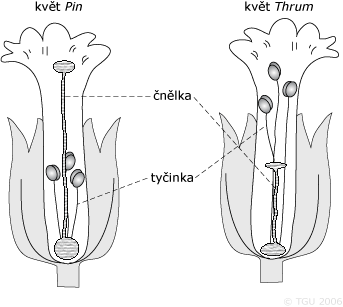
Příkladem může být polymorfizmus zvaný heterostylie u většiny druhů rodu prvosenek (Primula). Tento polymorfizmus má vztah k relativní délce čnělky (samičí orgán nesoucí bliznu) a tyčinek (samčí orgán nesoucí prašník) v květu. U prvosenek jsou dva typy květů: jeden známy jako pin, který má dlouhou čnělku a krátké tyčinky; druhý se označuj jako thrum, který má krátkou čnělku a dlouhé tyčinky. Negativní asortativní oplození je zde zprostředkováno opylujícím hmyzem (heterostyly), který umožňuje opylené obou typů květu navzájem (neboť samy o sobě se nemohou opylit) - opylovači, kteří sbírají pyl thrum na povrchu květu ho přenáší na bliznu pin; ale opylovači pracující uvnitř květu sbírají pyl pin a přenášejí ho na bliznu thrum. Navíc pylová zrna z květů pin se snadněji dostávají na receptor buněk blizny květů thrum; a pylová zrna z květů thrum klíčí lépe na bliznách květů pin než na svých vlastních. Rostliny pin se přednostně opylují rostlinami thrum.
Genetický efekt je opačný než u pozitivního asortativního oplození - zvyšuje se frekvence heterozygotů a snižuje homozygotů. To je způsobeno tím, že se častěji spolu páří homozygoti AA a aa navzájem, než by se očekávalo při náhodném oplození, protože nejsou fenotypově podobní. Genetický efekt se projevuje pouze na lokusech, které ovlivňují fenotypovou vlastnost důležitou pro oplození (a lokusy, které jsou s nimi ve vazbě). Negativní asortativní oplození může měnit frekvence alel pomocí procesu nazvanému negativní selekce závislá na frekvenci.

Aktualizováno: 03.02.2015
Aktualizováno: 03.02.2015