Selekce - výběr
Selekce (výběr) je hlavní evoluční síla a hlavní nástroj záměrného zlepšování kulturních rostlin a domestikovaných živočichů s cílem změny genového složení populace, tedy šlechtění. Selekce je síla, která zvyšuje nebo snižuje frekvenci alel v populaci.
Podstatou selekce je, že se na tvorbě nové generace podílí jen vybraná část rodičů (určité genotypy s žádoucím fenotypovým projevem - vyšší životnost, plodnost, adaptace, užitkovost), aby se rychleji rozmnožily žádoucí geny a nevýhodné geny byly eliminovány. Děje-li se tak působením přírodních faktorů, jedná se o přírodní selekci. Rozhoduje-li o výběru rodičů člověk, jedná se o selekci umělou. Lze rozlišit selekci pozitivní (zařazování rozmnožujících se jedinců) a negativní (vyřazování jedinců). Přírodní selekce působí ve všech ontogenetických fázích vývoje jedince. Člověk však při šlechtění zvířat selektuje hlavně dospělé jedince.
Selekce působí na:
- kvalitativní znaky - důsledky můžeme sledovat přímo pomocí změn ve frekvencích alel a genotypů,
- kvantitativní znaky - protože neznáme počet podílejících se genů, můžeme sledovat efektivnost selekce posunem průměrné fenotypové hodnoty ve směru selekce - studuje kvantitativní genetika.
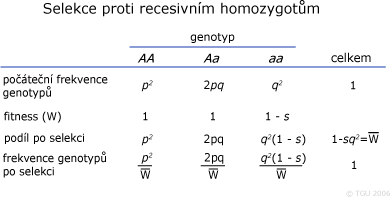
Fitness (W) vyjadřuje podíl potomků produkovaných jedním genotypem v porovnání s genotypem jiným – reprodukční způsobilost genotypu, adaptivní hodnota (někdy pojmenována jako selektivní hodnota - selective value). Pravděpodobnost, že nějaký fenotyp přežije a zanechá potomky je mírou jeho fitness. Jeho hodnota není stejná u všech jedinců v populaci (vliv prostředí a ostatních genů – genetické pozadí). Následný výklad předpokládá, že každý genotyp má svou vlastní hodnotu fitness. Přičemž fitness genotypů jsou konstantní a předpokládá se, že selekce se děje jen na základě různé přežitelnosti jedinců od zygot po dospělce (diploidní organizmy). Pokud je fitness každého genotypu rovna jeho pravděpodobnosti přežití, pak je každé fitness absolutním fitness, protože jeho hodnota je závislá na fitness ostatních genotypů. Obvykle však známe hodnotu životaschopnosti každého genotypu vztaženého relativně k ostatním vybraným genotypům jako standard k porovnání. Je-li hodnota fitness vyjádřena relativně k druhému genotypu, jedná se o relativní fitness. Relativní standardní fitness má hodnotu 1.
Síla selekce je vyjádřena hodnotou selekčního koeficientu (s). Jedná se o matematický rozdíl mezi hodnotou fitness jednoho genotypu a druhého genotypu. Oba koeficienty (W, s) jsou uváděny v relativních hodnotách v intervalu 0 až 1.
- s + W = 1
- W = 1 - s
- s = 1 - W
Jestliže všechny genotypy daného genu mají stejný počet potomků (AA:Aa:aa ~ 1:1:1), nepůsobí selekce na žádný genotyp a jejich fitness je W = 1.
Pravděpodobnost produkce gamet nesoucí recesivní alelu a je: P(aa) = f(aa)Waa = q2(1 - s).
Model selekce proti recesivním homozygotům
Známe-li frekvence alel (p, q) a hodnotu selekčních koeficientů genotypů, lze odhadnout změnu frekvencí alel a genotypů po jednotlivých generacích. Genotyp recesivně homozygotní je často spojen s negativním, škodlivým působením a proto se vůči němu vede negativní selekce, aby byl odstraněn z populace. Účinnost selekce proti recesivním homozygotům (saa = 1) závisí na výši frekvence recesivní alely.
| - frekvence q za 1 generaci | - frekvence p za 1 generaci | - frekvence q za n generací |
Úplná selekce (saa = 1) vůči genotypu recesivního homozygota v průběhu 1000 generací:
| Generace | frekvence p | frekvence q | p2 | 2pq | q2 |
|---|---|---|---|---|---|
| 0 | 0,5 | 0,5 | 0,25 | 0,50 | 0,25 |
| 1 | 0,67 | 0,33 | 0,45 | 0,44 | 0,12 |
| 2 | 0,75 | 0,25 | 0,56 | 0,38 | 0,06 |
| 3 | 0,80 | 0,20 | 0,64 | 0,32 | 0,04 |
| 4 | 0,833 | 0,167 | 0,694 | 0,278 | 0,028 |
| 5 | 0,857 | 0,143 | 0,734 | 0,245 | 0,020 |
| 10 | 0,917 | 0,083 | 0,841 | 0,152 | 0,007 |
| 40 | 0,976 | 0,024 | 0,953 | 0,047 | 0,001 |
| 70 | 0,986 | 0,014 | 0,972 | 0,028 | 0,0002 |
| 100 | 0,9902 | 0,0098 | 0,9805 | 0,0194 | 0,0001 |
| 200 | 0,9950 | 0,0050 | 0,9900 | 0,0100 | 0,00003 |
| 1000 | 0,9990 | 0,0010 | 0,9980 | 0,0020 | 0,000001 |
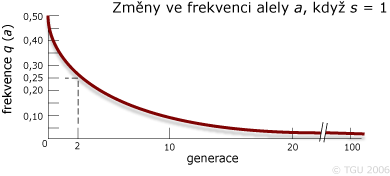
Změna četnosti alely a mezi generací rodičů a generací jejich potomků:
![]()
Přehled změn četnosti recesivní alely q za jednu generaci selekce při různých hodnotách intenzity selekce:
| Genotypy | Alely | ||||
|---|---|---|---|---|---|
| AA | Aa | aa | A | a | |
| frekvence | 0,36 | 0,48 | 0,16 | 0,60 | 0,40 |
| fitness | 1 | 0,95 | 0,70 | ||
| s | 0 | 0,05 | 0,30 | ||
| rodiče po selekci | |||||
| frekvence | 0,360 | 0,456 | 0,048 | ||
| skutečně se rozmnožující rodiče (po přepočtu na 100 %) | |||||
| frekvence | 0,4167 | 0,5278 | 0,0556 | 0,6808 | 0,3194 |
| potomci za 1 generaci selekce | |||||
| frekvence | 0,4632 | 0,4348 | 0,1020 | 0,6808 | 0,3194 |
Změna frekvence alel za jednu generaci selekce:
 p = + 0,0806
p = + 0,0806- q = - 0,0806
Průměrný fitness populace se jednoduše spočítá jako průměrný fitness všech jedinců v populaci: ![]() = p2 WAA + 2pq WAa + q2 Waa.
= p2 WAA + 2pq WAa + q2 Waa.
Změna ve frekvenci alely p: ![]()
Selekce proti různým genotypům
| waa = 0 | waa = 1 - s | |
| Selekce proti recesivním homozygotům | ||
| Selekce proti dominantní alele | : pro případ
WAA = 1-s WAa = 1-s Waa = 1 |
|
| Selekce proti oběma homozygotům | : pro případ
WAA = 1-s1 WAa = 1 Waa = 1-s2 |
|
Když s1 = s2 = 1, pak by již po 1 generaci selekce do genového fondu populace přispívali pouze heterozygoti ~ balancovaná letalita |
||
Změny četností alel mezi generacemi při selekci genotypů závisejí na:
- počáteční četnosti alel,
- fenotypovém projevu alel,
- hodnotě koeficientuv selekce,
- typu selekce,
- stupni dominance.
Na základě stručných charakteristik výše uvedených typu selekcí lze vyvodit následující obecně platné závěry:
- účinek selekce závisí na její intenzitě (vyjádřené hodnotou selekčního koeficientu) a počátečních genových četnostech,
- selekce proti dominantním nebo recesivním homozygotům (resp. proti odpovídajícím genotypům za nepřítomnosti dominance) vede postupně k rovnovážnému stavu, při kterém je příslušná alela z populace eliminována,
- selekce proti oběma homozygotům vede k ustavení rovnováhy při zachování dominantní i recesivní alely v populaci.

Aktualizováno: 31.10.2023