Genetické markery
Molekulárně-genetické analýzy kvantitativních znaků vedou k zjištění informací o dvou typech genetických lokusů (genetických markerů), využitelných v šlechtění.
- Prvním typem jsou kandidátní geny ~ přímé markery. Kandidátní geny mohou být charakterisovány dvěma typy polymorfismů:
- Funkční markery (Funkcional marker), což je vlastní příčinná mutace (nalezená např. na základě komparativního mapování)
- Přímé markery (Direct marker), což je polymorfismus DNA přímo v sekvenci genu
- Druhým typem jsou polymorfismy, které samy o sobě nemají vliv na projev znaku, ale jsou ve vazbě s QTL ~ nepřímé markery. Nepřímé markery je z hlediska praktického využití třeba rozdělit na dva typy:
- LD markery - markery které jsou s QTL ve vazbové nerovnováze (linkage disequilibrum markers)
- LE markery - markery, které jsou s QTL ve vazbové rovnováze (linkage equilibrum markers)
Z hlediska molekulární genetiky mohou být nepřímé genetické markery dvojího typu:
- vysoce variabilní mikrosatelitní sekvence (MS), nazývané také krátké tandemové repetice (STR; short tandem repeats).
- markery v kódujících i nekódujících sekvencích v intronech genů nebo mimo geny, v nichž se odhalil polymorfismus podmíněný záměnou (mutací) jedné baze v DNA. Tyto markery se označují zkratkou SNPs (sinle nucleotide polymorphism).
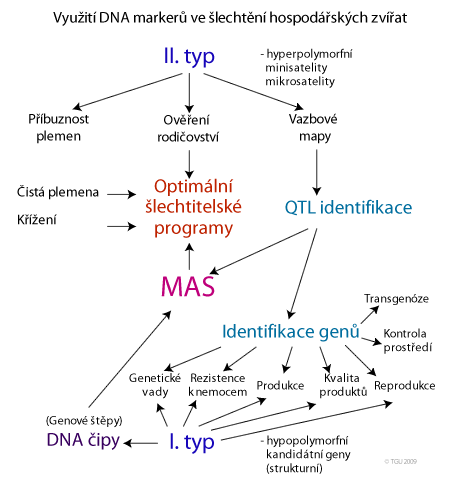
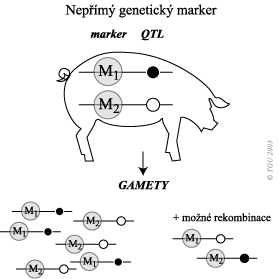
Schéma níže ukazuje princip závislosti informace o nepřímých genetických markerech a QTL. Můžeme identifikovat genotyp markeru (Mm), ale ne genotyp QTL (Qg). V případě že Q alela má pozitivní efekt a M alela je s ní spojována. Je vysoce pravděpodobné, že potomstvo získá od otce s alelou M také alelu Q, pokud nedojde k Crosing overu, a proto je alela M v selekci preferována. Z toho vyplývá, že využitelnost genetických markerů, je odvislá od těsnosti vazby mezi genetickým markerem a příčinnou mutací (existence vazbové nerovnováhy), či přesněji mutacemi v QTL. Čím je těsnost vazby vyšší, tím se zvyšuje i kvalita informace o genetickém markeru.
Vysvětlení genů velkého účinku pomocí genetických markerů
Přímé použití genů s určenou hodnotou může vést k rychlejšímu genetickému pokroku. To umožní genetické markery, které jsou definované jako úseky DNA bez funkce a bez vlivu na užitkovost. Jsou však snadno detekovatelné v laboratořích a tak lze říci, kterou alelu každého genetického markeru zvíře nese.
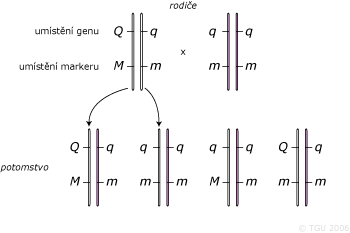
Nepřímý genetický marker je ve vazbě s QTL. Vyhledání těchto markerů vyžaduje dobře naplánovaný experiment. Vazba konkrétní alely markeru a QTL je neustále ohrožována rekombinacemi – funkcí vzdáleností mezi lokusy. Pro práci s nepřímými markery je potřeba provádět rodokmenovou analýzu a mít k dispozici změřené užitkové vlastnosti.
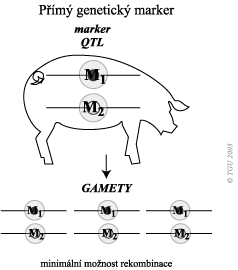
Je-li marker lokalizován přímo v QTL, pak již nejsou rekombinace problémem. Potřebujeme pouze změřit užitkové vlastnosti.
|
|
|
Proč se zajímat o genetické markery?
Nevýhody šlechtění na základě kvantitativní genetiky
- Nízké hodnoty koeficientu dědivosti. Např: plodnost a odolnost vůči chorobám.
- Vlastnosti, které nemohou být zjišťovány u všech jedinců. Jsou to vlastnosti, které se projevují jen u jednoho pohlaví nebo v dospělosti, tj. nikoliv u jedinců v raném věku.
- Některé vlastnosti nemohou být měřeny u živých zvířat, jako jsou ukazatele jatečné hodnoty a kvality masa.
- Vlastnosti mohou být zjištěny jen u omezeného počtu jedinců v testačním zařízení.
- Negativní korelace mezi vlastnostmi.
- Neaditivní genetické založení (dominance, epistáze). Odhad plemenné hodnoty a selekce se opírají o aditivně-genetickou proměnlivost a jsou opomíjeny intra a interlokuseová dědičnost, tj. dominance a epistaze.
- Při tvorbě syntetických populací nejsou často zohledňovány majorgeny, které mohou zapříčinit zlepšení užitkových vlastností.
- Neznáme genetické pozadí.
Některé výhody genetických markerů:
- Jsou zjistitelné v každém věku zvířete, nebo u embryí či ve spermatu.
- Markery pro znaky vyskytující se jen u jednoho pohlaví se dají zjistit i u druhého pohlaví.
- Pro znaky produkce a kvality masa které jsou hodnotitelné až po porážce a rozbourání, se mohou markery určit již u mladých zvířat.
- Varianty jednoho markeru mají mezi sebou kodominantní vztah, tzn. dají se přesně určit jednotlivé genotypy i heterozygotní, které se ve fenotypu neprojeví.
- Na základě znalostí genotypů markerů se mohou záměrně vybírat rodiče, provádět plemenitba a tak cíleně produkovat jatečná zvířata s určitými, ekonomicky výhodnými genotypy.
Využití markerů je příspěvkem pro zvýšení efektivnosti šlechtění v tomto směru:
- Přispívá ke zlepšení přesnosti odhadu plemenné hodnoty
- Umožňuje zvýšení intensity selekce
- Zkracuje genetický interval.
Jak využít informaci o GM?
Základní podmínkou pro využití genetických markerů je možnost stanovení genotypů v konkrétních genech u každého jedince. Následně je nezbytné určit jejich vliv na námi požadovaný znak. Suma těchto vlivů se dá označit jako MOLEKULÁRNÍ SKÓRE (molecular score). To pak může být využito jako selekční kriterium. Je samozřejmé, že využitelnost je vyšší u znaků u kterých známe celou podstatu genetického založení, vlivy jednotlivých genů a jejich interakcí s jinými geny či s prostředím. Pak můžeme využít znalost o sestavě alel nejen k selekci jedinců, ale například k záměrnému připařování, za účelem získání jedinců s konkrétní sestavou alel a tím i s konkrétními vlastnostmi.
Problémy při využití genetických markerů:
- První problém využití obou typů genetických markerů (tedy přímých i nepřímých) je v současné době nedostatek informací o tom které geny ovlivňují který znak. A proto je, vzhledem k možné složitosti interakcí ať již genetických či s prostředím, nemožné se se poléhat jen na informaci o genetických markerech. Do doby kdy budou informace o genetických markerech ucelenější a přesnější, je nutné využívat jak informaci o molekulárním skóre (které nám může dát například dobrou představu o tom jak daný jedince je odolný vůči určité chorobě, či zda jeho potomci budou mít dobrou kvalitu masa), tak i informace pocházejících z fenotypových hodnot (ty mají vyšší vypovídací hodnotu o celkovém genetickém založení a o jeho interakci s prostředím).
- Dalším problémem při využití nepřímých markerů je, že nesledujeme příčinnou mutaci, ale jen polymorfismus, který s ní je ve vazbě. Tato vazba však nemusí být stálá (otázka těsnosti vazby = vazbové rovnováhy a nerovnováhy), může dojít k rekombinacím (crosing–over), nebo se může v jiné populaci vyskytovat jiná vazbová fáze. Proto je důležité vybírat správné genetické markery pro danou populaci.
- Problémem společným pro přímé i nepřímé genetické markery, je že jejich vliv se stanovuje na základě znalostí o fenotypu. Zjištění jejich vlivu tedy může být zatíženo chybou vycházejících ze statistického zpracování, stejně jako z nepřesností při určování fenotypových hodnot.
- Testování DNA je stále relativné drahé v porovnání se zvýšením úrovně znaků.
- V případě že by došlo k vyselektování jedinců na homozygoty nemá daný marker další smysl. Příklad využití genu RYR1 (otázka genových zdrojů).
- Kvůli vysokému množství markerů je statisticky vysoká šance na chybné určení markerů majících vliv na daný znak. Již jen počet markerů by mohl vysvětlit více než 100% genetické variability u některých znaků.
- Pleiotropní a neaditivní efekty QTL. Proč se v populacích, které jsou dlouhodobě šlechtěny pro určitou užitkovost stále vyskytují v markerech alely, které mají negativní efekt na selektovanou vlastnost? To je velmi významná otázka. Jedna z možných odpovědí na tuto otázku je, že tyto alely mohou mít pozitivní efekt na fitnes a proto nebyly z populace vyselektovány na základě znalostí kvantitativní genetiky. Z toho vyplývá, že je nezbytné odhadnout pleiotropní a neaditivní efekty QTL na fitnes (případně další znaky) před jejich využitím v selekci. Jinak řečeno Markery mohou mít neznámé škodlivé nebo prospěšné efekty, proto by selekce neměla být prováděna do 100% fixace preferovaných alel v populaci.
- Ochrana objevů a investic. Firmy se snaží chránit svoje objevy pomocí patentů, což znemožňuje využití u jiných společností nebo států. Dalším problémem je otázka ochrany investic do MAS. Jestliže společnost investuje do MAS a získá pomocí ní jedince, kteří mají výrazně lepší užitkovost, jak chránit tento zisk před konkurencí. To je problémem především v chovu skotu, kde semeno od TOP býků, je k dispozici po celém světě. Oproti tomu u prasat a drůbeže není tato otázka příliš aktuální z důvodu firemních stád a hejn.
Aktualizováno: 16.01.2009