Biotechnologie zlepšující determinaci genetických hodnot zvířat
Odhad plemenných hodnot je v současné době prováděn analýzou fenotypu nebo fenotypové užitkovosti zvířat. Pokročilé statistické metody (BLUP, AM) se využívají pro oddělení genetických a prostřeďových efektů, při spojování zvířat genetickými příbuzenskými vztahy (Henderson, 1984). Fenotyp však nemůže být vždy zaznamenatelný z důvodů fyziologických (např. býci neprodukují mléko, kanci nemají selata, krávy rodí telata až po dvou letech života, …), některé vlastnosti nelze zjistit u živých zvířat (např. ukazatele jatečního těla, kvalita masa, …) nebo zaznamenávání fenotypu je chybné nebo nepřesné (snadnost telení). V posledních desetiletích došlo k hlubšímu poznání struktury DNA a působení určitých genů u jednotlivých druhů zvířat. Tyto výzkumy umožňují zkoumat, zda variabilita v DNA (polymorfizmus) určitých zvířat může být vázána k diferencím v produkčních nebo jiných ekonomických vlastností. Je zde však několik zásadních problémů vlastní celému genomu. Musí se určit funkční struktura chromatinu a pak určit v této struktuře funkční oblasti (geny, které se exprimují) a nefunkční oblasti (nám zatím s neznámou funkcí).
Hlavní výhody QTL/ETL ve šlechtitelských programech jsou:
- zvýšení přesnosti v selekci pomocí doplňkových informací přímo vztažených ke genotypu jedince
- možnost snížit generační interval přidáním nového selekčního období v životě zvířete – nejranější věk, protože QTL/ETL dovolují získat informace nezávislé na pohlaví a věku. Může také dojít ke zvýšení účinnosti introgrese nebo předpovídat heterozu pomocí genetické distance.
Metody využití genetických markerů v praktickém šlechtění
Metody využití genetických markerů, můžeme obecně rozdělit do dvou typů:
- Využití informací o fenotypu, rodokmenu a informací o markerech společně k vytvoření odhadu genetické hodnoty. Jedná se o rozšíření BLUP animal modelu, kterým je celkový genetický efekt rozdělen na dvě části, kdy první je efekt známých QTL a druhý je efekt neznámých genů (residual polygenes). Celková plemenná hodnota je pak dána součtem plemenné hodnoty dle QTL a efektem residuálních polygenů.
- Druhý je založen na dvoustupňovém výběru. V první fázi je informace o markerech využita k ranné selekci zvířat s žádoucími alelami. V druhé fázi jsou vyselektována zvířata testována na vlastní užitkovost a bez ohledu na informaci o markerech dále selektována.
Využití se liší podle toho o jaký typ markeru se jedná:
Gene Assisted Selection (GAS) Jak již bylo řečeno jde o využití polymorfismů (kandidátních genů) které jsou buď příčinnou mutací, nebo leží přímo v genu. Jejich využití v šlechtění je jednoduché, protože lze pro něj přímo určit vliv na ve šlechtění využívané znaky. Informace o markeru může být začleněna do odhadu plemenné hodnoty jako systematický efekt. Je pouze nutné určit vliv markeru v konkrétní populaci.
MAS využívající LD markery (LD-MAS) Jedná se o využití markerů, které jsou v vazbě s příčinnou mutací. Jejich využití se tedy příliš neliší od GAS. Při dlouhodobějším využívání je důležité sledovat zda nedojde ke snížení vypovídací hodnoty markeru z důvodu rekombinací. Důležité je zjistit vliv v každé populaci, a kterou chceme MAS aplikovat. V každé populaci může být jiná vazbová fáze s QTL nebo jiný vliv na základě rozdílného genetického pozadí.
MAS využívající LE markery (LE-MAS) Tyto marekery mají vypovídací schopnost pouze v populaci, ve které byly zjištěny, a tedy pouze v ní mohou být využívány. Vztah LE markeru k QTL může být odlišný v jednotlivých rodinách, a u různých generací. Proto je nutné průběžně v procesu MAS stanovovat vliv těchto markerů a to nejen u jedinců vybíraných do plemenitby, ale i u jejich příbuzných (z důvodu odhadu efektu QTL). Využití těchto markerů je vázáno dostupností (cenovou) molekulárních metod pro široké využití.
Konkrétní metody:
- program „sestavení genotypů“ (genotype building programes):
Jestliže známe QTL, a jestliže v různých liniích jsou obsaženy preferované alely, pak lze pomocí této metody získat jedince s novými výhodnějšími kombinacemi alel ve všech lokusech a QTL (například pro tvorbu finálních jedinců určených jen pro určitou produkci). Metoda je založena na molekulárním skóre, které je dáno genotypy v námi sledovaných lokusech (především nepřímými markery) a znalostí o jejich vazbové fázi. První fází je křížení dvou rodičovských linií, u kterých byla provedena testace a vybráni jedinci s homozygotní sestavou alel ve vybraných lokusech. Následuje několik generací ve kterých se vybírají k produkci další generace jedinci, kteří jsou homozygotní ve více než jednom lokusu. Tato metodika je vzhledem k svojí časové a metodické náročnosti využitelná spíše u rostlin než u zvířat.
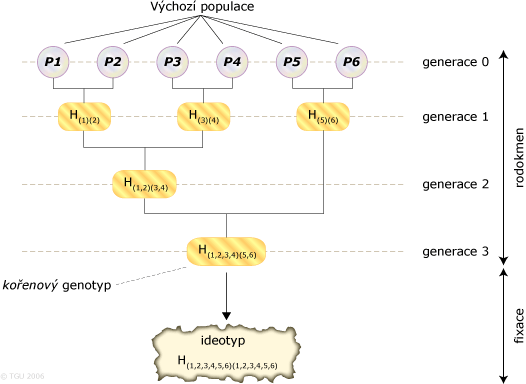
Servin et al. (2004) se ve své práci zabývali problematikou sestavení genetických pyramid, jejichž smyslem je shromáždit do jednoho genotypu alely, které byly zjištěny u většího počtu rodičovských populací. Předpokládali že je n lokusů, které jsou středem zájmu a to u n výchozích populací (P). U každé výchozí populace P se předpokládalo, že má homozygotní sestavu preferovaných alel ve vybraném lokusu i a ve všech ostatních n-1 lokusech má obě alely nepreferované. Smyslem je tedy získat ideální genotyp ve všech n lokusech. Tento genotyp můžeme označovat jako ideotyp. Postup vytvoření ideotypu lze označit jako "gene-pyramiding scheme". Každá výchozí populace je využita jen v jednom křížení v rámci pyramidy.
Celý proces lze rozdělit na dvě části:
- První část lze označit jako rodokmen (pedigree) a jejím cílem je kumulace jedné kopie alel všech cílových genů do jednoho kořenového genotypu (root genotype).
- Cílem druhé části nazvané fixace (fixacion step), je získat jedince homozygotní pro preferované alely ve všech vybraných lokusech (ideotyp).
Příklad genové pyramidy kumulující 6 cílových alel do jednoho ideotypu:
- program vnášení „genů“ (intorgression programmes; marker assisted introgression; MAI):
Jedná se o metodu, kdy do populace s nižší úrovní užitkovosti v určitém znaku vneseme a pomocí selekce zafixujeme alely v genu či genech zodpovědných za vývoj tohoto znaku z populace jiné. Základem je překřížení dvou populací donorské a recipientní. Následně pomocí selekce vybíráme jedince, kteří od donorské populace získali patřičné alely, ale v ostatních genech mají sestavu původní populace. Metoda je náročná na počet známých genů a alel v nich, tak aby spolu s alelami, které chceme od dárcovské populace přenést nedošlo i k vpravení jiných nechtěných alel.
Genetické markery zde mohou být použity dvojí cestou:
- Mohou pomoci identifikovat geny které by měly být "introgresovány".
- K selekci konkrétních genotypů.
- rekurentní selekční programy (recurrent selection programmes)
Pro jeden marker je možné molekulární skóre jedince, využitelné v opakované (rekurentní) selekci, určit jako odhad vztahu genetického markeru a fenotypu. Pro větší počet markerů je pak možno vytvořit jedno molekulární skóre (single molecular score) složené z dílčích molekulárních skóre jednotlivých genetických markerů (obdoba slekčního indexu). Kromě molekulárního skóre jsou k dispozici také informace o fenotypu jedince nebo jeho příbuzných. Možnost výběru jedince do plemenitby je tedy možná na základě kombinace těchto zdrojů informací o genetickém založení jedince. Existují tedy tři alternativy:
- Selekce jen na základě molekulárního skóre.
- Selekce na základě molekulárního skóre následovaná selekcí na základě fenotypu.
- Selekce na základě indexu kombinujícího informace o molekulárním skóre a fenotypu.
První varianta, tedy selekce jen na základě molekulárního skóre umožňuje v porovnání s dalšími dvěma variantami nejnižší genetické zlepšení. To by neplatilo pouze pokud by:
- informace o molekulárním skóre poskytovala ucelenou a úplnou informaci o genetickém založení znaků
- pokud by fenotypová data nebyla využitelná k selekci jedinců. Jako příklad může sloužit situace, kdy jeden či více členů rodiny musí být selektováno před možností zjištění fenotypových dat potřebných k odhadu plemenné hodnoty.
Pokud jsou k dispozici informace o molekulárním skóre i o fenotypu, pak pomocí kombinace těchto informaci je možné dosáhnout největšího šlechtitelského pokroku (druhá a třetí varianta). Při kombinaci těchto dvou zdrojů informací, je možné provádět selekci pomocí souhrnného selekčního indexu. Jeho sestavení zpracovali Lande and Thompson (1990). Index optimalizuje váhu molekulárního skóre a fenotypových informací, takže dochází k maximalizaci selekčního pokroku. Využití indexu je nejefektivnější pokud fenotypové informace jsou limitovány kvůli nízké heritabilitě, nebo nemožnosti získání informací o fenotypu u všech jedinců před vlastní selekcí. U QTL, k jejichž detekci jsou nezbytné informace o fenotypu, může u znaků s nízkou heritabilitou dojít k potížím při jejich detekci. Proto v některých případech může být využitelnost markerů vyšší u znaků s středním koeficientem heritability.
- křížení a hybridizace (crossbred or hybrid performance)
Křížení a hybridizace je hojně využívaná metoda založená na efektu heterose (využití neaditivních genetických efektů), kdy se předpokládá, že při křížení dvou divergentních linií se potomci vyznačují lepšími užitkovými hodnotami. Tento efekt je možné očekávat spíše u divergentních populací (v každé mohou být zafixovány jiné alely) než u geneticky příbuzných populací. Genetická vzdálenost mezi populacemi může být stanovena na základě rozdílů v frekvencích alel anonymních markerů napříč genomem. Zjištění vhodnosti populací k vzájemnému křížení na základě markerů je, na základě vyhodnocení této metody, využitelné spíše u populací, které mají společné předky, než u populací, které nejsou rodokmenově propojeny. U populací, které nevznikli ze stejných předků mohou existovat jiné vztahy mezi markerem a QTL (vazbová fáze, vazbová rovnováha).
MAS a současné šlechtitelské programy
Mnoho strategií využití markerů se snaží o začlenění informací o markerech do existujících šlechtitelských programů. Efektivní využití genetických markerů však může vyžadovat předělání stávajících programů tak aby bylo možné správně zjistit a odhadovat efekty QTL a následně co nejefektivněji využít výhod plynoucích z jejich znalosti. Je tedy nutné uzpůsobit MAS proces zjišťování a shromažďování fenotypových dat, způsob hodnocení zvířat způsob reprodukce apod. Jako příklad může posloužit jedna z výhod MAS a to zrychlení generačního intervalu pomocí zkrácení doby potřebné k výběru jedinců. To je však podmíněno využitím metod jako je oplodnění in vitro, embryotransfer, získávání oocytů od prepubertálních samic a dalších metod souvisejících s aplikací molekulárních technologií do procesu reprodukce.
Klíčovou roli v aplikaci genetických markerů do praxe je ekonomické zhodnocení. S MAS jsou spojeny výdaje na získání DNA, izolaci DNA až po výsledné zjištění genotypu a jeho vlivu na užitkové znaky. Využití markerů je tedy odpodstatněné tam, kde zisk s jejich použití je vyšší než zisk na základě šlechtění dle fenotypu a rodokmenových dat (využití odhadu plemené hodnoty pomocí BLUP-AM).
Marker-assisted selekce ke zlepšení užitkovosti v komerční chovech kříženců
Některé studie ukazují, že účinnost selekce čistokrevných plemen na zvýšení užitkovosti jejich komerčních kříženců potomků (CC - commercial crossbred) v běžných podmínkách je omezována nízkou genetickou korelací mezi užitkovostí čistokrevnými populacemi a komerčními kříženci. S tím se lze vypořádat začleněním fenotypových dat potomků kříženců pro selekci čistokrevných jedinců pomocí kombinované selekce kříženců a čistokrevných jedinců (CCPS). To může vést ke zvýšení stupně inbrídingu a navíc to vyžaduje komplexní systémy pro sběr fenotypových a rodokmenových dat u populací potomků kříženců. Používání odhadů efektů markerů na užitkovost kříženců pomocí MAS (CC-MAS) má své omezení. Model zahrnující informaci o markeru do selekčních strategií byl vyvinut na základě teorie selekčních indexů, která umožňuje předpovědět odpověď na selekci (genetický zistk) a stupeň inbrídingu při použití standardní deterministické selekční teorie. Pokud je genetická korelace mezi užitkovostí čistokrevných jedinců a kříženců rovna 0,7 pro šlechtitelský program představovaný finální otcovskou linii prasat, pak CC-MAS vykazuje značné zvýšení stupně odpovědi na selekci (genetický zisk) a snižuje stupeň inbrídingu v porovnání se selekcí čistokrevné populace a CCPS s 60 CC polosourozenci získaný pro každého čistokrevného kandidáta na selekci. Když je přesnost odhadu plemené hodnoty s pomocí markeru rovna 0,6, pak CC-MAS má za následek 34 a 10 % větší genetický zisk u užitkovosti CC v porovnání se selekcí u čistokrevných jedinců a CCPS. Korespondující stupně inbrídingu byly 1,4 % za generiaci pro CC-MAS, v porovnání s 2,1 % pro selekci čistokrevných jedinců a 3,0 % pro CCPS. Když byla přesnost odhadu plemené hodnoty pomocí markerů rovna 0,9, CC-MAS má za následek větší genetický zisk o 75 a 43 % než selekce čistokrevných jedinců a CCPS a snižuje stupeň inbrídingu o 1,0 % za generaci. Selekce na základě plemené hodnoty odhadnuté pomocí markeru odvozené z fenotypů čistokrevných jedinců má za následek podstatně menší genetický zisk u užitkovosti CC než v případě CC-MAS.
Závěrem lze říci, že efektivnost použití MAS vyžaduje odhady efektu užitkovosti komerčních kříženců (CC) a MAS založená na takovýchto odhadech umožňuje více efektivní selekci na užitkovost CC bez potřeby rozsáhlých rodokmenových záznamů, zatímco se snižuje stupeň inbrídingu. (Dekkers et al., 2007)
Výhled
Výhledově je pravděpodobné že bude stále vyšší důraz kladen na znaky spojené se zdravotní nezávadností živočišných produktů a kvality potravin pro člověka. Mnoho ze znaků spadajících do těchto oblastí vykazuje omezenou genetickou variabilitu a vysokou cenu stanovení fenotypu. Jako příklad může posloužit kvalita vepřového masa pro produkci šunky, resistence vůči parasitům, imunitní odezva. V rozvojových zemích pak mohou být QTL využity především pro resistenci k chorobám a k znakům adaptace.
Po stoletích se selektuje na komplexní vlastnosti pro zlepšení domestikovaných organizmů zcela podle fenotypu a rodokmenových dat (BLUP AM). Ačkoliv není důvod dokazovat až neočekávaně velké úspěchy touto metodou, věk genomiky nabízí možnost posunout selekci přímo ke genotypu. Je nutné říci, že gen velkého účinku rychle zvyšuje frekvenci při jednoduché fenotypové selekci. Takže v mnoha případech schopnost selektovat přímo na genotyp nemusí poskytnout až dramatické zvýšení genetického zisku. Evidentní případ, kde selekce na genotypy může potenciálně zvýšit zisk je, když můžeme selektovat jedince před fenotypovým projevem vlastnosti. BLUP AM předpověď plemenných hodnot pomocí informací o příbuznosti může provést stejné úlohy (ale ne ve všem, např. samci ze stejné rodiny sourozenců mají stejnou hodnotu BLUP, pokud mají stejné rodiče).
Je mnoho důvodů, proč by měl být fenotyp zahrnován do selekčních schémat. Jedním z nich je, že mutace tvoří novou variabilitu a selekce na fenotyp je snadný způsob jak uchovat tuto varianci. Dále selekce zaměřená na specifické genotypy může mít za následek mnohem větší snížení efektivní velikosti populace (Ne), než selekce založená jen na fenotypu. Snížení Ne snižuje akumulovanou mutační varianci, protože působí náhodný genetický drift a snižuje se genetický zisk v delším období.
Genomická revoluce je stejně důležitá jako jsou pokroky v reprodukčních technologií – embryotransfer, klonování, transgenní organizmy. Neméně důležitá je také revoluce v počítačích, která ovlivňuje využití poznatků z genomiky a budoucnost kvantitativní genetiky.
| Omezení pro kvantitativní genetiku | Příklad znaku | Zlepšení za využití molekulárních dat | Možná metoda šlechtění | Ekonomické zhodnocení využití molekulárních dat |
|---|---|---|---|---|
|
Fenotypová data jsou nedostatečná pro předpověď plemenné hodnoty (nízká heritabilita) |
Reprodukce |
Přesnější odhad plemenné hodnoty při využití informací o QTL |
Selekce na molekulární skóre a fenotyp |
Závislé na ceně stanovení markerů |
|
Zjištění fenotypu je obtížné nebo ekonomicky náročné |
Znaky spojené s onemocněními |
Markery jsou jednoduše stanovitelné a mohou být levnější ke stanovení |
Selekce na molekulární skóre |
Cena za zjištění fenotypu / cena za zjištění genotypu v markeru ? |
|
Projev fenotypu je odvislý od reprodukčního věku, dlouhý generační interval |
Reprodukce |
Molekulární skóre je zjistitelné v jakémkoliv věku = zkrácení generačního intervalu |
Kombinace selekce na molekulární skóre a na fenotyp předků |
Zrychlení použití lepšího genetického materiálu a tím zvýšení genetického zisku |
|
Genotyp je zjistitelný až po porážce jedince |
Kvalita masa |
Molekulární skóre je zjistitelné v jakémkoliv věku a z jakékoliv tkáně (chlupové cibulky apod.) |
Kombinace selekce na molekulární skóre a na genotyp příbuzných jedinců |
Značné zvýšení genetického zisku |
|
Znaky jsou pozorovatelné jen u jednoho pohlaví |
Mléčná užitkovost u skotu |
Molekulární skóre je zjistitelné v jakémkoliv věku, z jakékoliv tkáně a u obou pohlaví |
Kombinace selekce na molekulární skóre a na fenotyp. Například předselekce býků podle molekulárního skóre s následným testováním potomků. (progeny test) |
Otázka ceny předselekce a genetického zisku / normálnímu postupu šlechtění |
|
Genetický potenciál je skrytý v epistatických interakcích mezi QTL nebo vazbou QTL ve fázi repulsion |
Mnoho znaků |
Zjištění vazeb a vlivů na úrovni genů |
Kombinace selekce na molekulární skóre a na fenotyp. |
Obtížné odhadnout, může být vysoký, ale může být i malý, podle přesnosti odhadu efektu jednotlivých genů a jejich kombinací. |
|
Interakce genotyp-prostředí |
Mnoho znaků |
Předpověď interakce na genetické úrovni |
Kombinace selekce na molekulární skóre a na fenotyp. |
Neznámé, obtížné k předpovědi |
Aktualizováno: 16.01.2009