Nenáhodné oplození
- asortativní oplození
- inbríding
Inbríding
Definic slova inbrídingu je celá řada. Pokud se mezi sebou páří jedinci v populaci, kteří jsou úžeji příbuzní než by se očekávalo při náhodném páření, pak se tomu typu oplození říká inbríding (u zvířat ve šlechtění se označuje jako příbuzenská plemenitba).
U lidí se setkáme s nejužším stupněm inbrídingu na úrovni bratranec-sestřenice (first-cousin). Mnoho rostlin často podléhá samoopylení a některý hmyz často provádí oplození typu bratr-sestra. Inbríding nevyžaduje spojení úzce příbuzných jedinců. Určitá úroveň inbrídingu je nevyhnutelná v malých subpopulacích, protože členové malé subpopulace mají společné současné nebo vzdálené předky.
Genetická diferenciace mezi subpopulacemi, popisovaná hierarchickými F statistikami, může být vysvětlována typem efektu inbrídingu vyplývající z populační substruktury. Inbríding v přirozených populacích se může vyskytnout v případě malých populací, kde jsou potencionální rodiče s větší pravděpodobností více příbuzní než ve velkých.
Genotypové frekvence za inbrídingu
Hlavní efekt populační substruktury vlivem inbrídingu je snížení průměrné heterozygotnosti mezi subpopulacemi, relativně k heterozygotnosti očekávané za náhodného oplození v hypotetické velké populaci. Hlavní efekt inbrídingu je tvorba jedinců se sníženou heterozygotností, relativně k heterozygotnosti očekávané za náhodného oplození ve stejné subpopulaci. Snížení heterozygotnosti a tím pádem zvýšení homozygotnosti se týká všech lokusů (na rozdíl od asortativního oplození, kde se páří jedinci fenotypově podobní a mají tendenci sdílet stejné alely pouze na lokusech, které ovlivňují fenotypové vlastnosti ovlivňující páření). Důvodem, že inbríding ovlivňuje všechny lokusy stejně, je, že příbuzní jedinci jsou geneticky podobní podle společného původu a tedy je více pravděpodobné, že sdílejí stejné alely ve svých genomech než nepříbuzní jedinci.
Snížení heterozygotnosti v důsledku inbrídingu může být demonstrováno na příkladu samooplození (self-fertilisation). Uvažujme o samoopylující populaci rostlin, která obsahuje 1/4 AA, 1/2 Aa a 1/4 aa genotypy a populace je v těchto genotypech v Hardy-Weinberových rovnovážných poměrech. Protože každá rostlina podstupuje samoopylení, tak homozygotní genotypy vždy produkují jen homozygotní potomstvo a heterozygotní jedinci tvoří potomstvo 1/4 AA, 1/2 Aa a 1/4 aa. Po jedné generaci samoopylování vypadá potomstvo následně:
- AA: 1/4 x 1 + 1/2 x 1/4 = 3/8
- Aa: 1/2 x 1/2= 1/4
- aa: 1/4 x 1 + 1/2 x 1/4 = 3/8
Tyto genotypové frekvence nejsou v HW rovnováze. Došlo k výraznému snížení heterozygotů. V druhé generaci samoopylení jsou genotypové frekvence 7/16 AA, 2/16 Aa a 7/16 aa. Frekvence alel zůstává vždy stejná.
- v původní populaci: p = 1/4 + 1/2 x 1/2 = 1/2
- za jednu generaci samoopylení: p = 3/8 + 1/2 x 1/2= 1/2
- za dvě generace samoopylení: p = 7/16 + 1/2 x 2/16 = 1/2
V tabulce níže je příklad samooplození po čtyři generace, kde prvním jedincem je heterozygot v jednom genu.
| Generace | Genotypy | F | ||
|---|---|---|---|---|
| P1 |  |
0 | ||
| F1 | 0,250 | 0,500 | 0,250 | 1/2 |
| F2 | 0,375 | 0,250 | 0,375 | 3/4 |
| F3 | 0,437 | 0,125 | 0,437 | 7/8 |
| F4 | 0,468 | 0,063 | 0,468 | 15/16 |
| Fn | 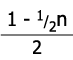 |
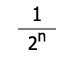 |
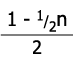 |
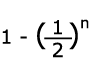 |
| 1/2 | 0 | 1/2 | 1 | |
Předpokladem je, že všechny genotypy mají stejnou pravděpodobnost přežití a rozmnožení. Je-li zde nějaká selekce proti nějakému genotypu, pak za inbrídingu se budou měnit i frekvence alel.
Efekt inbrídingu může být kvantifikován porovnáním poměru heterozygotních genotypů mezi inbredními organizmy s podílem heterozygotních genotypů očekávaných za náhodného oplození. Uvažujeme-li gen se dvěma alelami A a a, a frekvence heterozygotních genotypů v subpopulaci inbredních organizmů je HI. HWE frekvence heterozygotů by byla 2pq a je to označována symbolem H0. Efekt inbrídingu může být definován jako úměrné snížení v heterozygotnosti relativně k situaci za náhodného oplození. Tato hodnota je matematicky vyjádřena jako (H0- HI)/H0. Tento poměr se obvykle označuje symbolem F a nazývá koeficient inbrídingu.
![]()
F měří částečné snížení heterozygotnosti inbrední subpopulace ve vztahu k náhodně se pářící subpopulaci se stejnými frekvencemi alel. Protože H0 = 2pq, frekvence heterozygotních genotypů v inbrední subpopulaci může být zapsána v termínech F jako HI= H0 - H0F = H0(1 - F) = 2pq(1 - F).
Frekvence homozygotů AA v inbrední subpopulaci může být také zapsán pomocí F. Frekvence homozygotů je D a po dosazení do vzorce F = (H0- HI)/H0 získáme vztah D + HI/2 = p. Ale HI = 2pq(1 - F) a pak D = p - 2pq(1 - F)/2. Pokud platí p + q = 1, pak D = p - pq(1 - F) = p - pq + pqF = p(1-q) + pqF = p2 + pqF. Substitucí za q získáme: D = p2 + p(1-p)F = p2 + pF - p2F = p2(1-F) + pF.
Souhrn genotypových frekvencí v subpopulaci s koeficientem inbrídingu F:
- AA: p2(1-F) + pF = p2 + pqF
- Aa: 2pq(1 - F) = 2pq - 2pqF
- aa: q2(1-F) + qF = q2 + pqF
V inbrídingu je nedostatek heterozygotů roven 2pqF a přebytek každého homozygota roven polovině nedostatku heterozygotů. Pokud není v populaci inbríding (F = 0), pak jsou genotypové frekvence rovny rovnovážným frekvencím Hardy-Weinbergera; naopak s úplným inbrídingem (F = 1) má inbrední populace pouze genotypy AA a aa ve frekvencích p a q.
Vyskytuje-li se na lokusu více alel A1, A2, ..., An s frekvencemi p1, p2, ..., pn, pak v populaci s koeficientem inbrídingu F jsou frekvence genotypů:
- homozygotů AiAi: pi2(1-F) + piF
- heterozygotů AiAj: 2pipj(1 - F)
Koeficient inbrídingu jako pravděpodobnost
Interpretace koeficientu inbrídingu pomocí pravděpodobnosti je důležitá při výpočtu F z rodokmenu a předpokládá, že v v jednom inbredním organizmu jsou přítomny dvě alely jednoho genu. Organizmus je inbrední, pokud rodiče sdílí jednoho nebo více společných předků. Dvě alely přítomné v inbredním organizmu mohou být odvozeny ze stejné alely předka replikací DNA v jednom ze společných předků. V tomto případě se těmto případům říká, že jsou identické podle původu (identical by descent - IBD) a genotypy inbredního organizmu se nazývají autozygotní. Naopak, alely nemusí být replikami jedné alely předka a v tomto případě nejsou alely identické podle původu a genotypy se nazývají allozygotní.
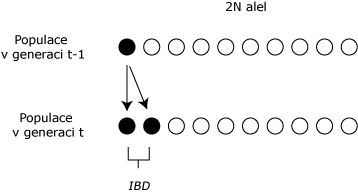
Charekterizace F pomocí pravděpodobnosti je, že dvě alely jednoho genu v inbredním organizmu jsou IBD (autozygotní). Obrázek níže zobrazuje jak je pojetí autozygotnosti a allozygotnosti vztaženo k homozygotnosti a heterozygotnosti. Dvě alely mohou být identické podle stavu (identical by state - IBS), což znamená, že alely mají stejnou sekvenci nukleotidů na DNA, bez ohledu na to, zda jsou IBD. Podle pojetí IBD, náleží alela podle původu po předcích a ne podle chemického složení. Dvě odlišné alely, které jsou IBS (dvě A1, nebo dvě A2 ), se mohou spojit při oplození do genotypu a tím vytvoří inbrední homozygotní organizmus a takový genotyp je allozygotní. Ačkoli heterozygotní genotypy musí být allozygotní (nebereme-li v úvahu mutace), homozygotní genotypy mohou být autozygotní i allozygotní.
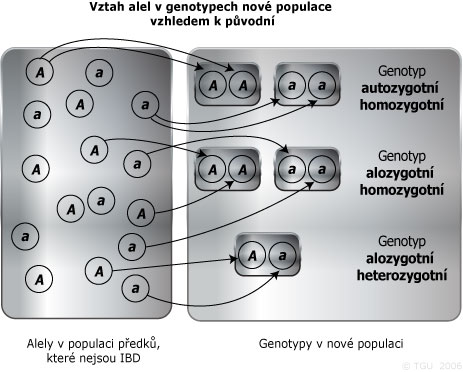
Alely v populaci lze rozdělit na:
- alozygotní alely s pravděpodobností (1-F) - náhodný inbriding je neovlivňuje tento gen, pravděpodobnost jakéhokoliv určitého genotypu v populaci je rovna pravděpodobnosti při náhodným oplozením,
- autozygotní alely s pravděpodobností F - jedinec musí být homozygotní a pak pravděpodobnost, že jedinec je homozygotní pro určitou alelu, je rovna četnosti této alely v populaci.
Genotypové frekvence za inbrídingu jsou souhrnně zobrazeny v obrázku níže. Obrazec je rozdělen vertikálně na dvě části korespondující s geny, jejichž alely zůstávají allozygotní navzdory působení inbrídingu a geny, jejichž alely jsou autozygotní z důvodu působení inbrídingu. Dělení je v podílu (1 - F) : F. V allozygotní části obrazce, horizontální panely odpovídají alozygotním genotypům AA, Aa a aa za genetické HW rovnováhy. V autozygotní části obrazce, horizontální panely odpovídají autozygotním genotypům AA a aa, které jsou v poměru p : q. Nevyskytují se zde heterozygotní genotypy, protože dvě alely přítomné v autozygotním lokusu jsou IBD.
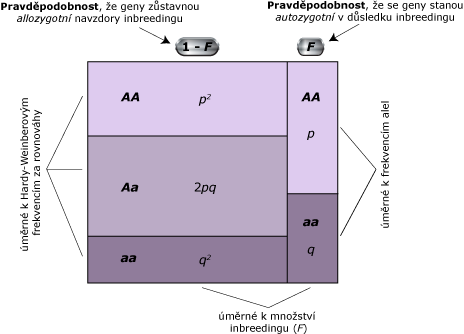
Genotypové frekvence jsou stejné jako ve vzorcích výše. Výsledky dokazují, že autozygotní definice F a heterozygotní definice F, ačkoliv jsou na pohled zcela odlišné, jsou ve skutečnosti obdobné.
| frekvence v populaci | ||||
|---|---|---|---|---|
| Genotyp | s koeficientem inbrídingu F | s F = 0 (náhodné oplození) |
s F = 1 (úplný inbríding) |
|
| AA | p2(1 - F) + | pF | p2 | p |
| Aa | 2pq(1 - F) | 2pq | 0 | |
| aa | q2(1 - F) + | qF | q2 | q |
| allozygotní geny |
autozygotní geny |
|||

Aktualizováno: 03.02.2015