
![]()
Příklad výpočtu c a p
|
U kukuřice (Zea mays) je pár alel C/c řídící zbarvení aleuronové vrstvy obilek ve vazbě s párem alel S/s, řídící tvar obilek. Mezi alelami jednotlivých genů platí vztah dominance. C – fialové zbarvení alleuronu, c - nezbarvený aleuron S – hladké obilky, s – svrasklé obilky Provede-li se klasický genetický pokus pro vznik hybridní generace a její testování zpětným křížením získáme všechny možné kombinace alel a fenotypů, jako by šlo o volnou kombinovatelnost. Oba geny jsou však na jednom chromozomu. Podíl jednotlivých fenotypů není roven poměru 1:1:1:1, jako při volné kombinovatelnosti. |
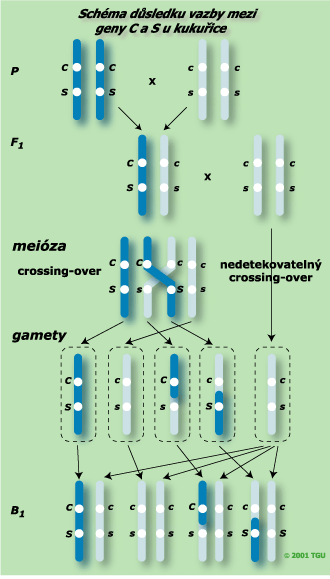
Křížené rodičovské komponenty jsou ve fázi cis. Dihybrid F1 generace tvoří nerekombinované gamety CS a cs s větším podílem a s menším podílem gamety rekombinované Cs a cS. Po zpětném křížení s recesivním homozygotem ccss byly zjištěny tyto hodnoty:
| Genotypy gamet F1 generace (CcSs) | CS | Cs | cS | cs |
| Genotypy gamet rodiče (ccss) | cs | cs | cs | cs |
| Genotypy obilek BC1 | CcSs | Ccss | ccSs | ccss |
| Fenotypy obilek BC1 | CS | Cs | cS | cs |
| Počet | 4032 | 149 | 152 | 4035 |
| Označení fenotypové třídy | a1 | a2 | a3 | a4 |
Batesonovo číslo:
c = (a1 + a4)/(a2 + a3) = (4 032 + 4 035)/(149 + 152) = 8 067/301 = 26,8
Gamety s rodičovskou sestavou vznikají 26,8krát častěji než rekombinované gamety.
Morganovo číslo:
p = (a2 + a3)/(a1 + a2 + a3 + a4) = (149 + 152)/(4 032 + 149 + 152 + 4035) = 301/8 368 = 0,035 M = 3,5 cM
Podíl rekombinovaných gamet je 3,5 % a vzdálenost mezi lokusy C a S je tedy 3,5 cM.
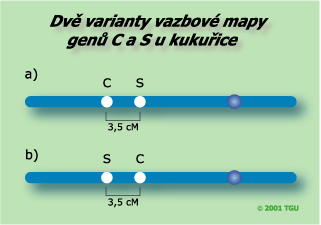
Při mapování dvou genů můžeme zjistit jejich vzájemnou vzdálenost, jako v tomto příkladě (3,5 cM mezi geny C a S). Nelze však určit v jakém pořadí se geny vyskytují od centromery. Nevíme tedy, zda jejich pořadí je S-C (a) nebo C-S (b). |
|
Konstrukce genetických map
Genetická mapa vyjadřuje:
- vazbou skupinu,
- symboly mutantních genů,
- vzdálenost genů v centimorganech z jednoho konce chromozomu, který je považovaný za nulový bod,
- označení centromery.
|
|
Genetická mapa 6. chromozomu u skotu a prasete. |
Genetická mapa 1. chromozomu octomilka. |


Cílem genetického mapování je určit pořadí genů a jejich vzdálenosti na chromozomech. Kromě genetických (rekombinačních) map, využívající rekombinační frekvence a popisující vzdálenosti genů v cM, se konstruují i fyzické mapy založené na sekvencování a jednotkami jsou přímo nukleotidy (lidský genom byl osekvencován na jaře roku 2001). Jejich informace se sjednocují v nejpřesnějším mapování genů ve spojení s cytogenetickými technikami.
Pro genetické mapovaní je výhodnější použít tříbodový test, kdy současně sledujeme dílčí vazbové vztahy (rekombinační frekvence) 3 různých genů, např. A, B, C.
|
x |
| Gamety F1 | Genotypy zygot | Počet jedinců | Počet jedinců v % |
nerekombinované (rodičovské) |
|
580 592 celkem 1172 |
80,94 |
jednoduchý c.-o. mezi B a C
|
|
45 40 celkem 85 |
5,87 |
jednoduchý c.-o. mezi A a B
|
|
89 94 celkem 183 |
12,64 |
dvojitý c.-o. |
|
3 5 celkem 8 |
0,55 |
Celkem |
1448 |
100 % |
Postup:
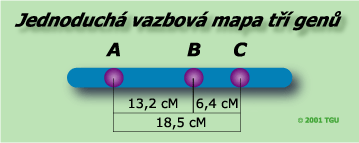
- Určíme rodičovské genotypy. Jsou to vždy ty, s největší frekvencí. Zde se jedná o genotypy ABC a abc.
- Určíme pořadí genů. Vycházíme ze znalosti dvojitého crossing-overu, který určíme z nejnižší frekvence genotypů - zde AbC a aBc. Je nutné si uvědomit, že dvojitý crossing-over přesunuje prostřední alelu mezi nesesterskými chromatidami. Můžeme vidět, že gen B musí být uprostřed, protože recesivní alela b je nyní na stejném chromozomu jako alely A a C a dominantní alela B je na stejném chromozomu jako recesivní alely a a c. Pořadí genů na chromozomu je ABC.
- Určíme vazbové vzdálenosti mezi geny. Určíme vazbové vzdálenosti mezi geny A-C a C-B. Vazba se vypočítá jako podíl celkového počtu rekombinantních gamet k celkovému počtu gamet (~ Morganovo číslo p). Je třeba vzít v úvahu dvojitý crossing-over. Do obou výpočtů vzdálenosti se proto začlení jeho hodnota.
- A-B vzdálenost: (89+94+3+5)/1448 = 0,1319 ~ 13,19 cM
- B-C vzdálenost: (45+40+3+5)/1448 = 0,0642 ~ 6,42 cM
- Jestliže je správné konstatování, že překřížení je funkcí vzdálenosti mezi geny, pak můžeme stanovit mezi geny A a C, jako součet dvou frekvencí jednoduchých c.-o.: 13,19 + 6,42 = 19,61 cM. V daném případě však celkový počet jednotlivých c.o. mezi geny A a C činí (89+94+45+40)/1448 = 0,1851 ~ 18,51 cM. Vzdálenost mezi geny A a C vyjádřena součtem jednotlivých c.o. je větší o 1,1 cM než celá vzdálenost A-C.
- Určení vzdálenosti mezi geny A a C: 18,51 + 2x0,55 = 19,61 cM. Zdvojení procenta dvojitých c.o. je nutné proto, že každý dvojitý c.o. vzniká na základě dvou nezávislých jednoduchých zlomů ve dvou bodech.
- Nakreslení mapy.
| |
Interference a koeficient koincidence
Mapování genů je tím přesnější, čím jsou vzájemně v silnější vazbě a čím méně se uplatňuje tzv. interference. Do asi 20 - 25 cM je mapová vzdálenost shodná s rekombinační frekvencí. Nad 20 - 25 cM se vzdálenosti "prodlužují" (vzrůstá počet vícenásobných rekombinací) a sčítání dílčích mapových vzdáleností je více nepřesné a nad 0,35 cM nespolehlivé.
Interference (I) se měří mezi crossing-overy v dané oblasti chromozomu. Vyskytne-li se dvojitý crossing-over, lze uvažovat o interferenci. Dané specifické rekombinační poměry ve dvou sousedních chromozomových vzdáleností, pak poměr dvojitých crossing-overů je v této oblasti roven součinu jednoduchých crossing-overů: (0,1319 x 0,0642)*100 = 0,847 % dvojitých rekombinací. V našem případě by se jednalo o 12,3 dvojitých rekombinantů (1448 * 0,00847). Ve skutečnosti jich bylo odhaleno jen 8.
K měření interference je nutné nejdříve vypočítat koeficient koincidence (c.o.c.), který je dán poměrem pozorovaných k očekávaným dvojitým crossing-overům (2c.o.). Interference je pak rovna 1- c.o.c.
![]()
Pro náš případ je hodnota interference 33 % [(1 - 8/12)*100].
Vazbová nerovnováha
Při neúplné vazbě dvou genů na homologních chromozomech se předpokládá jejich rovnoměrné rozložení v populaci - frekvence všech 4 kombinací ve stejném poměru (AB, Ab, aB, ab). Často dochází k odchylkám a nerovnoměrnému rozložení - určité kombinace se vyskytují častěji než jiné. Tomuto stavu nerovnoměrné frekvenci alel vázaných genů se říká vazbová nerovnováha. Vysvětlení možných příčin:
- populace se vyvíjí v izolaci s inbridingem a náhodným genetickým driftem,
- asociace více alel je dána jejich evoluční výhodností,
- od vzniku alel neuplynula delší doba, aby dosáhly rovnováhy.
Využití vazbové nerovnováhy se v současné době hledá v mapování lokusů kvantitativních vlastností (QTL) pomocí genetických markerů a její možné využití ve šlechtění.
![]()
![]()
![]()