
![]()
Jestliže vybereme náhodně dva geny vázané na jednom chromozomu, je vysoce pravděpodobné, že budou tak blízko jeden druhému podél chromozomu, že představují úplnou vazbu. Při úplné vazbě nedochází při meióze mezi geny ke crossing-overu. Pokud budeme křížit dva náhodně vybrané geny vázané na jednom chromozomu, budou téměř vždy produkovat určitý podíl potomstva, vzniklého z rekombinantních gamet. Jejich podíl je proměnlivý a závisí na vzdálenosti mezi dvěma geny na chromozomu. Pokud jsou genové páry jednoho chromozomu umístěny dále jeden od druhého, pak hovoříme o neúplné (částečné) vazbě. Při neúplné vazbě dochází při meióze k výměně genetického materiálu mezi nesesterskými chromatidami homologů. Obecně se tento proces nazývá genetická rekombinace.
Meiotický základ rekombinací
Ve stádiu pachytene může dojít ke zlomu chromatid s následnou fůzí sesterských nebo nesesterských chromatid. Při reciprokých výměnách mezi sesterskými chromatidami nedochází ke genetické variabilitě a výměna není detekovatelná. Klíčová je výměna u nesesterských chromatid.
- Při crossing-overu nedochází ke ztrátě nebo k přidání chromatinu.
- Překříží se vždy pouze dvě chromatidy.
- Může se vyskytnout i vícenásobný crossing-over mezi nesesterskými chromatidami.
- Může nastat jakákoliv crossoverová konfigurace a výsledek může být velmi odlišný od původní kombinace alel na chromatidě.
- Ke crossing-overu dochází až po replikaci chromozomu.
Mechanizmus crossing-overu
Při crossing-overu dochází k fyzické výměně mezi molekulami DNA dvou homologních chromozomů. Zásadní význam má chiazma pozorované během profáze I meiózy a zlom s opětným sjednocením. Sledují se dvě teorie založené na tvorbě chiazma, ale zcela různými způsoby.
Klasická teorieCrossing-over je výsledkem nahodilé fyzické výměny v chiazma. Překřížení v chiazma je zodpovědné za crossing-over a zcela jasně jej předchází. Tato teorie předpovídá, že ke crossing-overu dochází po stádiu diplotene, ale před oddělením chromozomů v anafázi I. |
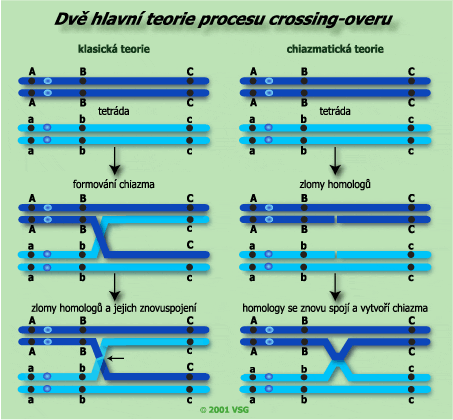 |
Chiazmatická teoriepředpovídá, že crossing-over předchází formování chiazma a dochází k němu v raném stádiu pachytene profáze I. Jako výsledek je, že chiazmata jsou tvořena místy genetické výměny, takže jsou důsledkem crossing-overu a ve stádiu diplotene dochází k projevu chromozomové výměny. |
Typy crossing-overů
|
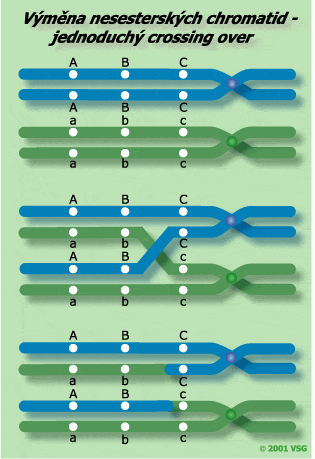
Rekombinační procesy se dějí náhodně po celé délce tetrády. Čím blíže jsou umístěné lokusy podél chromozomu, tím méně pravděpodobné je, že mezi nimi proběhne jednoduchý crossing-over. Naopak čím vzdálenější lokusy, tím větší pravděpodobnost crossing-overu. Když dochází k jednoduchému crossing-overu mezi dvěma nesesterskými chromatidami, druhé dvě chromatidy tetrády jsou neovlivněny a vstupují do gamet nezměněny. I kdyby vždy (u 100 %) došlo k jednoduchému crossing-overu mezi dvěma vázanými geny, mohou být rekombinace pozorovány postupně u 50 procent potenciálně tvořených gamet. |
|
Vícenásobné crossing-overy
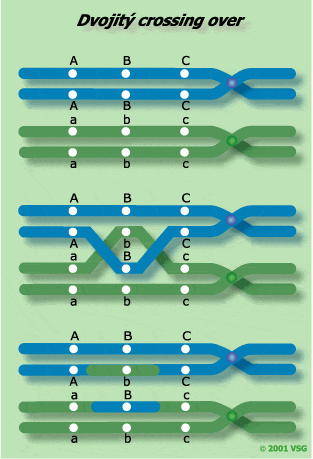
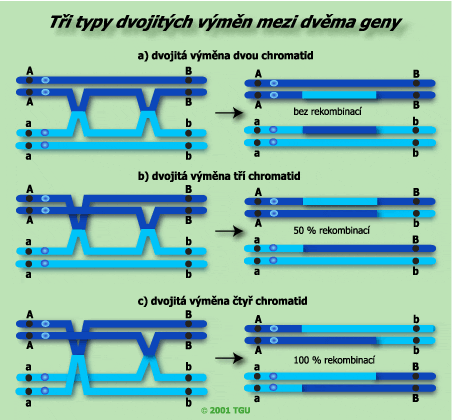
Dochází také k tomu, že v jedné tetrádě se vymění genetický materiál mezi dvěma, třemi i více místy nesesterských chromatid, jako důsledek více crossing-overů. Dvojité výměny genetického materiálu vyplývají z dvojitých crossing-overů. Pro jejich studium je třeba sledovat tři vázané geny. |
|
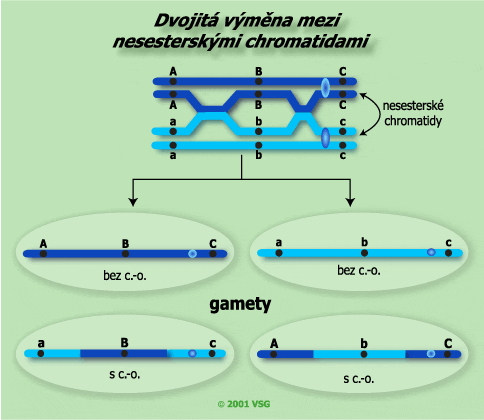
Výsledek dvojité výměny probíhající mezi nesesterskými chromatidami. Protože k výměně došlo pouze mezi dvěma chromatidami, tvoří se dvě necrosované gamety (rodičovské) a dvě dvojitě crosované (rekombinantní). |
|
Zde jsou tři možné způsoby dvojitého crossing-overu. |
![]()
![]()
![]()
![]()