Model DNA podle Watsona a Cricka
Úvod:

Za vhodných podmínek se DNA může vyrovnat do vlákna, aby se vytvořila správná struktura. DNA vlákna rozptylují X paprsky (Röntgenové záření) a tvoří na ploše charakteristické ohyby (difrakce). Metoda krystalografie X paprsky (zkoumáním struktury krystalů organických sloučenin metodou rentgenové krystalografie) naznačila třídimenzionální strukturu DNA. Následnou fotografii difrakce DNA pomocí X paprsků získali Rosalinda Franklinová a Maurice Wilkins v roce 1952. Křížový vzor odrazu X paprsku naznačuje, že DNA je helikální - má dva spirálovité řetězce.
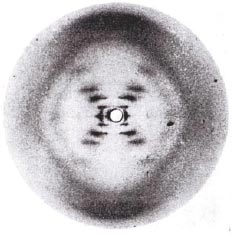
[Maddox B. The double helix and the "wronged heroine". Nature, 2003, vol. 421: 407-408.]
Tyto poznatky využili ve svém výzkumu James Watson a Francis Crick, z kterého vzešel platný model DNA jako dvojitá šroubovice dvou antiparalelně spojených nukleotidových řetězců, které spojují navzájem dusíkaté báze vodíkovými vazbami. Struktura DNA byla publikována v roce 1953.
Zatímco Watson a Crick a Wilkinson za svůj objev získali Nobelovu cenu v roce 1962, Franklinová se toho nedožila (1920 - 1957), ačkoli její práce výrazně napomohla poznání struktury DNA (zacházení s X paprsky má své důsledky).
Je zajímavé, že u objevu století, zásadním objevu pro genetiku, nestáli genetikové, ale fyzici a biochemikové.
Nukleotidy jsou základní stavební bloky DNA
DNA je dlouhý polymer složený z podjednotek známých jako nukleotidy. Každý nukleotid se skládá z cukru deoxyribózy, fosfátu a dusíkaté báze. Znalost detailní chemické struktury těchto složek a způsob jejich kombinování hraje důležitou roli v modelu, který sestavili Watson a Crick.
Na první uhlík deoxyribózy se kovalentně napojuje příslušná báze a tvoří nukleosid.
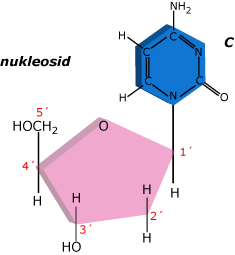
Připojení fosfátové skupiny na 5´uhlík cukru se tvoří kompletní nukleotid.
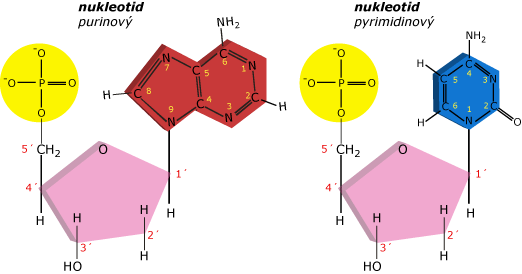
Polarita DNA řetězce složeného z mnoha nukleotidů
DNA řetězec složený z mnoha nukleotidů má polaritu - směr. Fosfodiesterové vazby vždy tvoří kovalentní spojení mezi 3´ uhlíkem jednoho nukleosidu a 5´ uhlíkem následujícího nukleosidu. Souhlasná orientace nukleotidů dává řetězci obecný směr, tak že dva konce jednoho řetězce jsou chemicky rozdílné.
Na 5´konci má cukr terminálního nukleotidu volný 5´ uhlík (ve smyslu, že není vázán na jiný nukleotid). V závislosti na tom, jak je DNA syntetizována, 5´uhlík nukleotidu na 5´konci může nést buď hydroxylovou nebo fosfátovou skupinu. Na 3´konci řetězce je 3´uhlík posledního nukleotidu volný. Po celém řetězci je polarita 5´-3´ zachovávána od nukleotidu k nukleotidu. Tato konvence se nazývá směr 5´-3´, který jde ve směru zleva doprava.
Směrová sekvence nukleotidových bází může nést informaci
Informace může být zakódována pouze v sekvenci symbolů, jejichž pořadí se liší podle zprávy, která má být zapsána. Bez sekvenční variability není potenciál pro zapsání informace. Protože hlavní kostra DNA je dvojice cukr-fosfát je chemicky idnetická pro každý nukleotid v řetězci DNA, rozdílnost může přinést pouze variabilita dusíkaté báze. Jestliže DNA nese genetickou informaci, tato informace musí být zapsána ve variabilitě sekvenci A, G, T a C bazí. Různá kombinace čtyř bazí v dlouhé sekvenci nukleotidů může kódovat informaci pro tvorbu organizmu.
Komplementarita bazí je strukturní podstavou genetické funkce
Watson-Crickova struktura byla založena na porozumění a interpretaci všech chemických a fyzikálních dat, dostupných v té době. Své zjištění publikovali ve vědeckém časopisu Nature v dubnu roku 1953.
Jakmile objevili strukturu, okamžitě z ní vyplynula funkce DNA, tzn. jak je genetická informace zapsána a jak se přenáší z generace do generace. Z výsledků rentgenové kristalografie vyplynulo, že v molekule DNA musí být více než jeden řetězec nukleotidů.
Nukleotidy vykazují komplementaritu
Když je DNA složena z více polynukleotidových řetězců, jaká síla je drží při sobě? Na to odpověděl svým výzkumem Erwin Chargaff, když porovnával nukleotidové složení DNA z různých organizmů. Ačkoliv mezi jednotlivými druhy jsou velké rozdíly v množství jednotlivých bází, poměr A:T byl vždy 1:1 a poměr G:C byl také 1:1.
|
% bazí |
poměry |
|||||
|
organizmus |
A |
T |
G |
C |
A:T |
G:C |
|
Escherichia coli |
26,0 |
23,9 |
24,9 |
25,2 |
1,09 |
0,99 |
|
Kvasinka |
31,3 |
32,9 |
18,7 |
17,1 |
0,95 |
1,09 |
|
Caenorhabditis elegans |
31,2 |
29,1 |
19,3 |
20,5 |
1,07 |
0,96 |
|
Arabidopsis thaliana |
29,1 |
29,7 |
20,5 |
20,7 |
0,98 |
0,99 |
|
Drosophila melanogaster |
27,3 |
27,6 |
22,5 |
22,5 |
0,99 |
1,00 |
|
včela |
34,4 |
33,0 |
16,2 |
16,4 |
1,04 |
0,99 |
|
myš |
29,2 |
29,4 |
21,7 |
19,7 |
0,99 |
1,10 |
|
člověk |
30,7 |
31,2 |
19,3 |
18,8 |
0,98 |
1,03 |
Vysvětlení Chargafových poměrů je možné jen, když budeme uvažovat o chemické afinitě mezi A a T a mezi G a C. Porovnáním bazí došel Watson k závěru, že mezi purinovou a pyrimidinovou bází jsou vodíkové vazby: slabá elektrostatická vazba mezi atomy vodíku.
Komplementární párování bází
Adenin na jednom řetězci může tvořit dvě vodíkové vazby s tyminem druhého řetězce. Guanin jednoho řetězce tvoří tři vodíkové vazby s cytozinem druhého řetězce DNA. Velikost a tvar A-T a G-C párů bazí jsou podobné a zaujímají stejný prostor mezi oběma polydeoxyribonukleotidovými řetězci helixu DNA.
Sekundární struktura DNA - Watson-Crickův model
DNA je dlouhý polymer složený z podjednotek známých jako nukleotidy. Každý nukleotid se skládá z cukru deoxyribózy, fosfátu a dusíkaté báze. Znalost detailní chemické struktury těchto složek a způsob jejich kombinace hraje významnou roli v modelu DNA vytvořeného Watsonem a Crickem.
DNA řetězec složený z mnoha nukleotidů má polaritu - směr. Fosfodiesterové vazby vždy tvoří kovalentní spojení mezi 3´ uhlíkem jednoho nukleosidu a 5´ uhlíkem následujícího nukleosidu. Souhlasná orientace nukleotidů dává řetězci obecný směr, tak že dva konce jednoho řetězce jsou chemicky rozdílné.
Na 5´konci má cukr terminálního nukleotidu volný 5´ uhlík (ve smyslu, že není vázán na jiný nukleotid). V závislosti na tom, jak je DNA syntetizována, 5´uhlík nukleotidu na 5´konci může nést buď hydroxylovou, nebo fosfátovou skupinu. Na 3´konci řetězce je 3´uhlík posledního nukleotidu volný. Po celém řetězci je polarita 5´-3´ zachovávána od nukleotidu k nukleotidu. Tato konvence se nazývá směr 5´-3´, který jde ve směru zleva doprava.
Směrová sekvence bazí může nést informaci
Informace může být zakódována pouze v sekvenci symbolů, jejichž pořadí se liší podle zprávy, která má být zapsána. Bez sekvenční variability není potenciál pro zapsání informace. Protože hlavní kostra DNA je dvojice cukr-fosfát je chemicky identická pro každý nukleotid v řetězci DNA, rozdílnost může přinést pouze variabilita dusíkaté báze. Jestliže DNA nese genetickou informaci, tato informace musí být zapsána ve variabilitě sekvenci A, G, T a C bazí. Různá kombinace čtyř bazí v dlouhé sekvenci nukleotidů může kódovat informaci pro tvorbu organizmu.
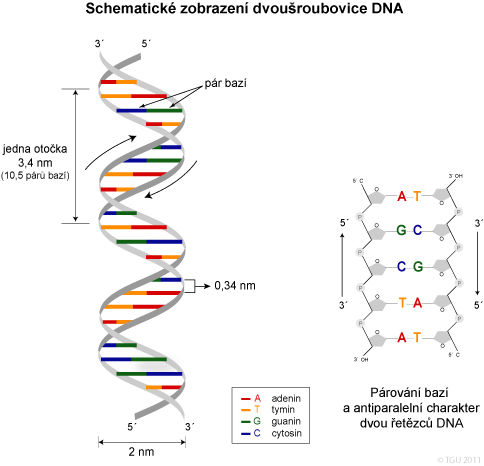
Závěr:
- difrakce DNA X paprsky naznačila helikální strukturu DNA,
- nukleotidy jsou základní stavební kameny DNA,
- dvojitý helix obsahuje dva antiparalelní řetězce, které se spojují na základě komplementarity bazí,
- dvojitý helix může tvořit alternativní formy,
- DNA struktura je základem genetické funkce.