
![]()
![]()
|
Je-li populace v genetické rovnováze, je stabilizovaná bez dalšího vývoje - evoluční stagnace. V reálných populacích zvířat a rostlin, kdy nejsou splňovány výše zmíněné podmínky rovnováhy, je H.-W. genetická rovnováha neustále porušována a zároveň populace neustále směřují k obnovení rovnováhy. Genetická rovnováha je porušována řadou vnějších a vnitřních faktorů. Přirozené populace jsou dynamické, mění svou velikost a strukturu - evoluční vývoj.
Faktory evolučních změn
- systematické (nenáhodné, soustavné) - lze určit směr a velikost změny v četnosti alel a genotypů; opakované mutace, jednosměrné migrace, dlouhodobý selekční tlak
- stochastické
(náhodné) - nelze určit směr, ale jen rozsah změn četnosti alel a
genotypů; náhodný (genetický) drift, náhodné změny v migraci, ve
směru a intenzitě selekce
Selekce
Selekce (výběr) je hlavní evoluční síla a hlavní nástroj záměrného zlepšování kulturních rostlin a domestikovaných živočichů s cílem změny genového složení populace, tedy šlechtění. Selekce je síla, která zvyšuje nebo snižuje frekvenci alel v populaci.
Podstatou selekce je, že se na tvorbě nové generace podílí jen vybraná část rodičů (určité genotypy s žádoucím fenotypovým projevem - vyšší životnost, plodnost, adaptace, užitkovost), aby se rychleji rozmnožily žádoucí geny a nevýhodné geny byly eliminovány. Děje-li se tak působením přírodních faktorů, jedná se o přírodní selekci. Rozhoduje-li o výběru rodičů člověk, jedná se o selekci umělou. Lze rozlišit selekci pozitivní (zařazování rozmnožujících se jedinců) a negativní (vyřazování jedinců). Selekce působí ve všech ontogenetických fázích vývoje jedince. Člověk však při šlechtění zvířat selektuje hlavně dospělé jedince.
Selekce působí na:
- kvalitativní znaky - můžeme sledovat přímo změny ve frekvencích alel a genotypů.
- kvantitativní znaky - protože neznáme počet podílejících se genů, můžeme sledovat efektivnost selekce posunem průměrné fenotypové hodnoty ve směru selekce - studuje kvantitativní genetika.
Fitness (W) vyjadřuje podíl potomků produkovaných jedním genotypem v porovnání s genotypem jiným – reprodukční způsobilost genotypu, adaptivní hodnota (někdy pojmenována jako selektivní hodnota - selective value). Pravděpodobnost, že nějaký fenotyp přežije a zanechá potomky je mírou jeho fitness. Jeho hodnota není stejná u všech jedinců v populaci (vliv prostředí a ostatních genů – genetické pozadí).
Síla selekce je vyjádřena hodnotou selekčního koeficientu (s). Jedná se o matematický rozdíl mezi hodnotou fitness jednoho genotypu a druhého genotypu.
Oba koeficienty (W, s) jsou uváděny v relativních hodnotách v intervalu 0 až 1.
Jestliže všechny genotypy daného genu mají stejný počet potomků (AA:Aa:aa ~ 1:1:1), nepůsobí selekce na žádný genotyp a jejich fitness je W = 1.
|
Pravděpodobnost produkce gamet nesoucí recesivní alelu a:
![]()
Selekce proti recesivním homozygotům
| Známe-li frekvence alel (p, q) a hodnotu selekčních koeficientů genotypů, lze odhadnout změnu frekvencí alel a genotypů po jednotlivých generacích. Genotyp recesivně homozygotní je často spojen s negativním, škodlivým působením a proto se vůči němu vede negativní selekce, aby byl odstraněn z populace. Účinnost selekce proti recesivním homozygotům (saa = 1) závisí na frekvenci recesivní alely. |
|
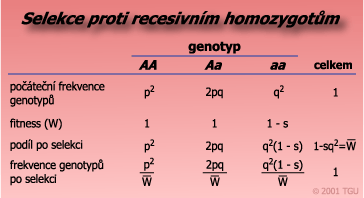
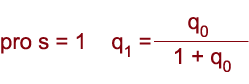 |
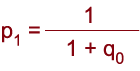 |
||
- za n generací: |
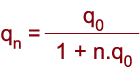 |
||
Úplná selekce (s = 1) vůči genotypu recesivního homozygota v průběhu 1000 generací.
| Generace | frekvence p | frekvence q | p2 | 2pq | q2 |
| 0 | 0,5 | 0,5 | 0,25 | 0,50 | 0,25 |
| 1 | 0,67 | 0,33 | 0,45 | 0,44 | 0,12 |
| 2 | 0,75 | 0,25 | 0,56 | 0,38 | 0,06 |
| 3 | 0,80 | 0,20 | 0,64 | 0,32 | 0,04 |
| 4 | 0,833 | 0,167 | 0,694 | 0,278 | 0,028 |
| 5 | 0,857 | 0,143 | 0,734 | 0,245 | 0,020 |
| 10 | 0,917 | 0,083 | 0,841 | 0,152 | 0,007 |
| 40 | 0,976 | 0,024 | 0,953 | 0,047 | 0,001 |
| 70 | 0,986 | 0,014 | 0,972 | 0,028 | 0,0002 |
| 100 | 0,9902 | 0,0098 | 0,9805 | 0,0194 | 0,0001 |
| 200 | 0,9950 | 0,0050 | 0,9900 | 0,0100 | 0,00003 |
| 1000 | 0,9990 | 0,0010 | 0,9980 | 0,0020 | 0,000001 |
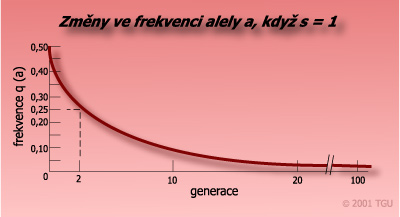
Změna četnosti alely a mezi generací rodičů a generací jejich potomků:
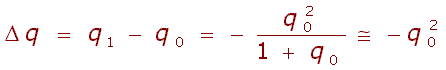
|
Přehled změn četnosti recesivní alely q za jednu generaci selekce při různých hodnotách intenzity selekce:
Genotypy |
Alely | ||||
| AA | Aa | aa | A | a | |
| frekvence | 0,36 | 0,48 | 0,16 | 0,60 | 0,40 |
| fitness | 1 | 0,95 | 0,30 | ||
| s | 0 | 0,05 | 0,70 | ||
| rodiče po selekci | |||||
| frekvence | 0,360 | 0,456 | 0,048 | ||
| skutečně se rozmnožující rodiče (po přepočtu na 100 %) | |||||
| frekvence | 0,4167 | 0,5278 | 0,0556 | 0,6808 | 0,3194 |
| potomci za 1 generaci selekce | |||||
| frekvence | 0,4632 | 0,4348 | 0,1020 | 0,6808 | 0,3194 |
![]()
![]()
![]() -
průměrný fitness populace
-
průměrný fitness populace
![]()
![]()
![]()
![]()