
![]()
Duplicitní faktory (dimerie) zodpovídají za několik od sebe odlišných typů interakcí. Mají však společný základní charakter, kterým se odlišují od vzájemného doplňování (komplementarity), vztah nadřízenosti a podřízenosti (epistáze - hypostáze) či vzájemné potlačování (kompenzace), že totiž určitý efekt je vyvolaný více genovými páry, z kterých však každý už sám je schopný tento efekt způsobit.
Identické dominantní či aktivní alely multiplicitních genů odpovídají za biosyntézu identických konečných produktů (jednoho a téhož znaku), avšak kvalitativně rozdílnými způsoby (každá z dominantní - aktivní alely zúčastněných lokusů schopen vyvolat znak, avšak intenzita, kvalita projevu závisí na vzájemných vztazích duplicitních genů). Rozlišují se duplicitní faktory nekumulativní s dominancí, kumulativní s dominancí a kumulativní bez dominance.
![]()
Duplicitní faktory nekumulativní s dominancí
– při této interakci je každá dominantní alela schopná vyvolat příslušný znak, přičemž však přítomnost většího počtu těchto vloh se neprojevuje na intenzitě znaku. Při nekumulativní multiplicitě konečný produkt podmíněný kteroukoliv dominantní alelou tohoto druhu, tedy vzniklý jednou ze všech možných biosyntetických cest, vyvolá již plný fenotypový projev znaku.
Příklad:
genotypy A1A1A2A2, A1A1A2a2, A1a1a2A2, A1a1a2a2 atd. mají jeden fenotyp a genotyp a1a1a2a2 má druhý fenotyp.
| P | A1A1a2a2 x a1a1A2A2 | |
| GP | A1a2 | a1A2 |
| F1 | A1a1A2a2 | |
| GF1 | A1A2, A1a2, a1A2, a1a2 |
|
| F2 | A1A2 | A1a2 | a1A2 | a1a2 |
| A1A2 | A1A1A2A2 | A1A1A2a2 | A1a1A2A2 | A1a1A2a2 |
| A1a2 | A1A1A2a2 | A1A1a2a2 | A1a1A2a2 | A1a1a2a2 |
| a1A2 | A1a1A2A2 | A1a1A2a2 | a1a1A2A2 | a1a1A2a2 |
| a1a2 | A1a1A2a2 | A1a1a2a2 | a1a1A2a2 | a1a1a2a2 |
nekumulativní s dominancí
genotypový štěpný poměr – 1 : 2 : 1 : 2 : 4 : 2 : 1 : 2 : 1
fenotypový štěpný poměr – 15 : 1
Testovací křížení F1 x recesivní homozygot: A1a1A2a2 x a1a1a2a2
| FB | A1A2 | A1a2 | a1A2 | a1a2 |
| a1a2 | A1a1A2a2 | A1a1a2a2 | a1a1A2a2 | a1a1a2a2 |
fenotypový štěpný poměr 3 : 1
Touto interakcí se dědí tvar šešule u kokošky pastuší tobolky (Capsela bursa pastoralis), u živočichů se tak dědí opeření běháků u slepic.
![]()
Duplicitní faktory kumulativní dominancí
– je genotyp s oběma faktory dominantně přítomnými, ať v homozygotním nebo heterozygotním stavu, fenotypově odlišný od genotypů, kde je z obou párů zastoupený vždy jen jeden. Ve vyjádření fenotypu se sčítá (kumuluje) efekt aktivních alel lokusů. Fenotypově se bude lišit i genotypy homozygotně recesivní. Při kumulativních multiplicitách je intenzita fenotypu odstupňována podle počtu dominantních či aktivních alel a tím i podle počtu biosyntetických cest, po nich příslušné reakce proběhly až ke konečnému produktu.
Příklad: V případě dvou lokusů o dvou alelách:
- genotyp A1-A2- (černý fenotyp)
- genotyp A1-a2a2, a1a1A2- (šedý fenotyp)
- genotyp a1a1a2a2 (bílý fenotyp)
| P | A1A1a2a2 x a1a1A2A2 | |
| GP | A1a2 | a1A2 |
| F1 | A1a1A2a2 | |
| GF1 | A1A2, A1a2, a1A2, a1a2 |
|
| F2 | A1A2 | A1a2 | a1A2 | a1a2 |
| A1A2 | A1A1A2A2 | A1A1A2a2 | A1a1A2A2 | A1a1A2a2 |
| A1a2 | A1A1A2a2 | A1A1a2a2 | A1a1A2a2 | A1a1a2a2 |
| a1A2 | A1a1A2A2 | A1a1A2a2 | a1a1A2A2 | a1a1A2a2 |
| a1a2 | A1a1A2a2 | A1a1a2a2 | a1a1A2a2 | a1a1a2a2 |
kumulativní s dominancí
genotypový štěpný poměr – 1 : 2 : 1 : 2 : 4 : 2 : 1 : 2 : 1
fenotypový štěpný poměr – 9 : 6 : 1
Testovací křížení F1 x recesivní homozygot: A1a1A2a2 x a1a1a2a2
| FB | A1A2 | A1a2 | a1A2 | a1a2 |
| a1a2 | A1a1A2a2 | A1a1a2a2 | a1a1A2a2 | a1a1a2a2 |
fenotypový štěpný poměr 1 : 2 : 1
Příkladem může být barva zrna u ječmene (rostliny s hnědočervenými zrny mají alespoň 1 aktivní alelu ze dvou genů; recesivní homozygoti mají zrna světlá; rostliny, mající alespoň 1 aktivní alelu v obou genech, mají zesílený fenotyp - tmavohnědá zrna).
![]()
Duplicitní faktory kumulativní bez dominance
– tento typ interakce se vyznačuje tím, že mezi alelami jednotlivých párů je buď poměr neúplné dominance, nebo není vůbec možno hovořit o dominanci, ale je možné rozeznat mezi dvěma různými alelami jednu aktivní, která má vliv na fenotyp a druhou neutrální, která nemá vliv na zvýraznění příslušného fenotypu. Kombinací dvou duplicitních kumulativních genů bez dominance dostaneme tedy pět různých fenotypů, pět různých stupňů intenzity daného znaku (maximální, tříčtvrtinový, poloviční, čtvrtinový a nulový.
Příklad:
- genotyp A1A1A2A2 – fenotyp 1 (maximální)
- genotyp A1A1a2A2, A1a1A2A2 - fenotyp 2 (tříčtvrtinový)
- genotyp A1A1a2a2, a1a1A2A2, A1a1A2a2 – fenotyp 3 (poloviční)
- genotyp A1a1a2a2, a1a1A2a2 – fenotyp 4 (čtvrtinový)
- genotyp a1a1a2a2 – fenotyp 5 (nulový)
| P | A1A1a2a2 x a1a1A2A2 | |
| GP | A1a2 | a1A2 |
| F1 | A1a1A2a2 | |
| GF1 | A1A2, A1a2, a1A2, a1a2 |
|
| F2 | A1A2 | A1a2 | a1A2 | a1a2 |
| A1A2 | A1A1A2A2 | A1A1A2a2 | A1a1A2A2 | A1a1A2a2 |
| A1a2 | A1A1A2a2 | A1A1a2a2 | A1a1A2a2 | A1a1a2a2 |
| a1A2 | A1a1A2A2 | A1a1A2a2 | a1a1A2A2 | a1a1A2a2 |
| a1a2 | A1a1A2a2 | A1a1a2a2 | a1a1A2a2 | a1a1a2a2 |
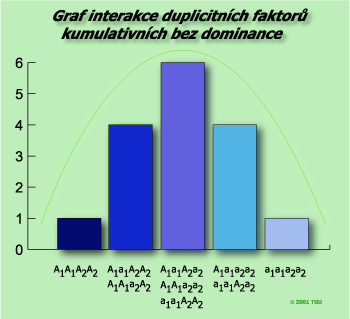
kumulativní bez dominance
genotypový štěpný poměr – 1 : 2 : 1 : 2 : 4 : 2 : 1 : 2 : 1
fenotypový štěpný poměr – 1 : 4 : 6 : 4 : 1
Testovací křížení F1 x recesivní homozygot:A1a1A2a2 x a1a1a2a2
| FB | A1A2 | A1a2 | a1A2 | a1a2 |
| a1a2 | A1a1A2a2 | A1a1a2a2 | a1a1A2a2 | a1a1a2a2 |
fenotypový štěpný poměr 1 : 2 : 1
Tímto typem interakce se řídí celá řada fyziologických znaků, např. ranost u ovsa nebo rýže (4 páry genů).
| Z grafu je patrné, že aditivní duplicitní interakce je přechod k dědičnosti kvantitativních znaků s kontinuální proměnlivostí (polygenní dědičnost), při které může jít o aditivitu nebo o kumulaci. Trihybrid štěpí v poměru 1:6:15:20:15:6:1 (další poměry můžete odvozovat z Pascalova trojúhelníka). |
|
![]()
![]()
![]()