Replikace DNA jako kopírování genetické informace pro přenos do další generace
Na konci článku Watsona a Cricka z roku 1953, v kterém popsali strukturu modelu dvojitého helixu DNA, se nachází věta: "It has not escaped our notice that the specific pairing we have postulated immediately suggests a possible copying mechanism for the genetic material." přeloženo: "Neušlo naší pozornosti, že specifické párování může umožňovat kopírovací mechanizmus pro genetický materiál."
Komplementární vztah mezi bázemi, tedy specifické párování bazí, předpokládá možný kopírovací mechanizmus pro genetický materiál. Kopírování genetické informace předchází jejímu přenosu pomocí chromozomů z jedné generace do druhé na základě meiózy. Je také základem pro duplikování chromozomů před každou mitózou, která zabezpečuje, že všechny dceřiné buňky vyvíjejícího se organizmu získají kompletní kopii genetické informace.
Teprve objevením molekulární struktury DNA vedla biology k přijetí závěrů Oswalda T. Averyho, že DNA a ne proteiny je nosičem genetické informace. Studium replikace DNA byla však teprve na začátku. Replikace jednoduché molekuly, jakou je DNA, je komplexní, vícekrokový proces, zahrnující mnoho rozličných enzymů. Replikace lineárního eukaryotického chromozomu je ještě komplexnější, než replikace kruhového prokaryotního chromozomu.
Genetická kontinuita mezi rodičovskými a dceřinými buňkami je umožněna semikonzervativní replikací DNA, kterou předpokládali již Watson a Crick. Byl navrhován také proces konzervativní replikace. Proces replikace, v kterém každý řetězec dvojitého helixu DNA slouží jako templát pro syntézu nového vlákna, je v principu jednoduchý. Jedná se o tvorbu identických kopií molekul nukleových kyselin, což zajišťuje přenos genetické informace z DNA do DNA nebo z RNA do RNA. Syntéza DNA je komplexní proces, řízený velkým množstvím enzymů a dalších molekul s jediným cílem – s velkou přesností polymerizovat nukleotidy do polynukleotidových řetězců.
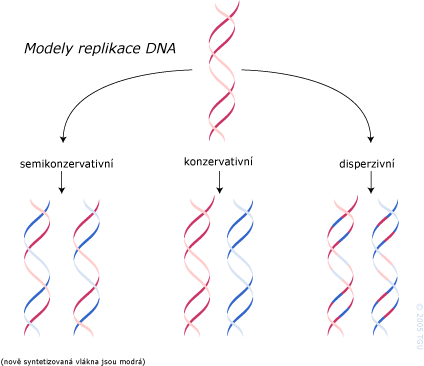
Semikonzervativní model replikace
Při procesu replikace se podle Watsona a Cricka dvojitý helix rozvine a odhalí báze v obou řetězcích DNA. Každý ze dvou oddělených řetězců slouží jako templát, molekulární matrice pro syntézu nového řetězce.
Nový řetězec se tvoří na základě komplementarity bazí, jako protikladný řetězec vůči původnímu rodičovskému řetězci. Tzn., že A na jedné pozici v původním řetězci dává podnět k připojení komplementární báze T na korespondující pozici nově formovaného dceřiného řetězce. Podobně se vůči T v původním řetězci přidá A v novém řetězci, vůči G se přidá C a vůči C přisedne G. Toto párování bazí, které určuje nukleotidovou sekvenci nového řetězce, se nazývá komplementární párování bazí.
Když vhodná báze vytvoří vodíkovou vazbu se svým komplementem, enzym spojí nukleotid k předcházejícímu nukleotidu fosfodiesterovou vazbou (popřípadě navázáním celého nového nukleotidového řetězce do kontinuálního řetězce). Tento mechanizmus komplementárního párování bazí, následovaný spojením po sobě následujících nukleotidů, vytváří „dceřiné“ dvojité helixy, kde každý obsahuje jeden původní řetězec DNA a jeden kompletně nový.
Z toho důvodu se tomuto typu duplikace dvojitého helixu říká semikonzervativní replikace: kopírování, kdy jeden řetězec je konzervativní pocházející z rodičovské molekuly DNA (sloužil jako templát) a druhý je nově syntetizovaný.
Existují další dva možné modely replikace: konzervativní a disperzivní. Tyto alternativy jsou však nevyhovující, protože nenaznačují mechanizmus, kterým by docházelo k replikaci genetické informace v sekvenci bazí a nevysvětlují experimentální důkazy, svědčící o semikonzervativní replikaci (důkaz podali ve svém experimentu Meselson a Stahl).
Experiment Meselsona a Stahla - důkaz semikonzervativní replikace
V roce 1958 Matthew Meselson a Franklin Stahl (California Institute of Technology,
Nechali růst bakterii E. Coli po několik generací na médiu, v kterém byl dusík s normálním izotopem 14N (tato kultura sloužila jako kontrola). Druhou kulturu nechali růst v médiu s dusíkem, který měl těžký izotop 15N. Po několik generací byly v DNA veškeré atomy dusíku značeny 15N (zabudovali se do dusíkatých bazí DNA – A, T, G, C). Buňky z těchto kultur byly přeneseny do nového média s dusíkem 14N. Nově syntetizované DNA řetězce by měly obsahovat lehčí izotopy dusíku.
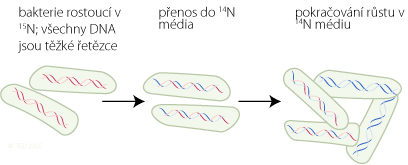
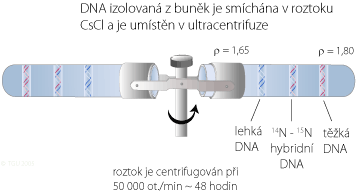
Meselson a Stahl izolovali DNA z buněk rostoucích v rozdílných izotopových kulturách a pak vystavili tyto DNA vzorky gradientní hustotní centrifugaci (analytická technika, kterou vyvinuli). Protože DNA obsahující 15N je hustší než DNA obsahující 14N, objeví se v centrifugační zkumavce dva proužky. Čistá 15N DNA bude tvořit proužky níže ke dnu centrifugační zkumavky, než DNA s 14N.
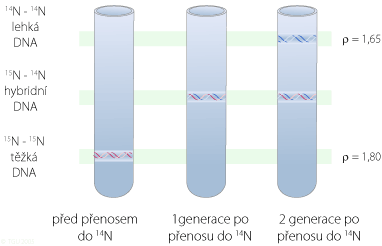
Výsledky experimentu byly ve shodě pouze s principem semikonzervativní replikace, kterou navrhli Watson a Crick. V první generaci po přenosu z 15N do 14N média, jeden ze dvou řetězců každé dceřiné DNA nesl těžké izotopy dusíku a druhý nově syntetizovaný řetězec obsahoval jen lehké izotopy dusíku 14N. Proužek nacházející se mezi proužky pro 14N DNA a 15N DNA představuje jeho hybridní izotop. V druhé generaci po přenosu měla polovina molekul DNA jeden řetězec 15N a druhý 14N, zatímco zbývající polovina DNA nesla dva 14N řetězce. Dva pozorovatelné proužky (jeden hybridní a jeden z čistých 14N) představují tuto směs.
Nyní se předpokládá, že semikonzervativní replikace DNA je téměř univerzální.
Meselson, M. & Stahl, F. W. (1958) Proc. Natl. Acad. Sci. USA 44, 671–682.