Translace - tvorba proteinů
Úvod k translaci
Prokaryotické i eukaryotické buňky používají stejný základní genetický kód, aby se přeložila sekvence nukleotidů v mRNA do sekvence aminokyselin v odpovídajícím polypeptidu. Translace se děje na ribozomech, které koordinují pohyb transferových RNA nesoucích specifické aminokyseliny s genetickými instrukcemi obsaženými v mRNA.
Struktura a funkce tRNA
Protože není žádná zřejmá chemická podobnost nebo afinita mezi triplety nukleotidů mRNA (kodony) a příslušnými aminokyselinami, je nutný nějaký prostředník, transferová RNA (tRNA). Tato tRNA slouží jako přechodová molekula, která zprostředkuje přenos informace z nukleové kyseliny do proteinu.
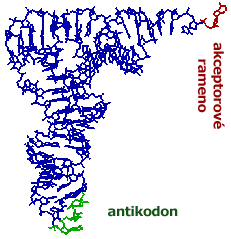
Molekuly tRNA mají antikodon na jednom konci a aminokyselinu na druhém
Molekuly tRNA jsou krátké, jednořetězcové RNA molekuly v délce 74 – 95 nukleotidů. Ve struktuře se objevují i modifikované báze vzniklé chemickými změnami nukleotidů A, G, C a U (např. ribotymidin, 5-metylcytidin, inosin, 7-metylguanosin, ...). Každá tRNA nese jednu konkrétní aminokyselinu a v buňce musí být minimálně jedna tRNA pro každou z 20 aminokyselin. Označení tRNA je podle toho, kterou aminokyselinu nese – tRNAGly nese aminokyselinu glycin.
Nukleotidová sekvence tRNA tvoří primární strukturu. Krátké komplementární úseky v tomto řetězci můžou tvořit páry bází a tak vytváří charakteristické tvary jetelových lístků – sekundární struktura. Složením do prostoru se vytváří terciární struktura, která vypadá jako písmeno L. Místa pro antikodon a pro připojení aminokyseliny jsou na opačných koncích tRNA.

Párování bazí kodon-antikodon na ribozomu se děje antiparalelně – např. je-li antikodon na tRNA 3´ CGU 5´, je komplementární kodon na mRNA 3´GCA 5´, specifikující aminokyselinu alanin.
Terciární struktura tRNA je dána dalšími vazbami mezi komplementárními páry bází na řetězci.
Aminoacyl-tRNA syntetáza katalyzuje esterifikaci korespondující aminokyseliny s tRNA
Enzym amonoacyl-tRNA syntetáza nejdříve použije energii z ATP hydrolýzou a aktivuje aminokyselinu a tvoří aminoacyladenylát – karboxylová skupina aminokyseliny reaguje s ATP. Aminoacyladenylát se váže na enzym, dokud se nesetká s molekulou tRNA, specifickou pro tuto amonoacyl-tRNA syntetázu. Enzym pak přenese aktivovanou aminokyselinu na 3´ konec tRNA (na 3´OH konec nebo na 2´OH konec ribózy) a vytvoří se aminoacyl~tRNA. Mezi aminokyselinou a tRNA se vytvoří makroergická vazba. Tím je dokončena aktivace.
Párování mezi kodonem mRNA a antikodonem tRNA
Aminokyselina sama o sobě nehraje žádnou roli v určení, kde se začlení do rostoucího polypeptidického řetězce. To je specifikováno výhradně nukleovými kyselinami, mezi kodonem mRNA a antikodonem tRNA. Např. tRNA s antikodonem pro cystein (3´ACA 5´), ale nesla by místo toho aminokyselinu alanin, byl by alanin začleněn do polypeptidu kdykoliv, kdy by v mRNA byl kodon 5´UGU 3´.
Ribozomy - továrny na proteosyntézu
Ribozomy jsou komplexní struktury složené z RNA a proteinů
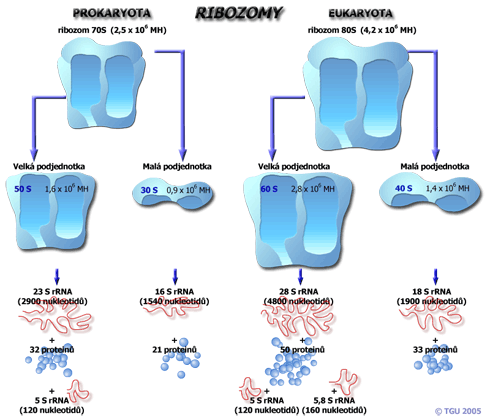
Ribozomy mají dvě podjednotky, každá složená z ribozomální RNA (rRNA) a různých proteinů. Je rozdíl mezi ribozomy prokaryotů a eukaryotů. Před translací jsou obě jednotky oddělené v cytoplazmě. Krátce po začátku translace se spojují do kompletního ribozomu. Eukaryotické ribozomy mají více komponent než prokaryotické, jsou to však pořád dvě podjednotky.
Ribozomy mají mnoho funkcí při proteosyntéze:
- rozpoznávají startovací sekvenci v mRNA,
- pomáhají zabezpečit přesnost interpretace genetického kódu, tím, že stabilizují interakci mezi tRNA a mRNA (rozpoznání kodon-antikodon by bylo velmi slabé),
- doplňují enzymatickou aktivitu, když spojují aminokyseliny v rostoucím polypeptidickém řetězci,
- při pohybu 5´> 3´podél mRNA, vystavují kodony mRNA v sekvenci a zabezpečují tak, že se lineárně přidávají aminokyseliny,
- pomáhají ukončit syntézu polypeptidu, když rozdělují mRNA a protein od sebe a uvolnění proteinu samotného.
Různé části ribozomu mají různé funkce
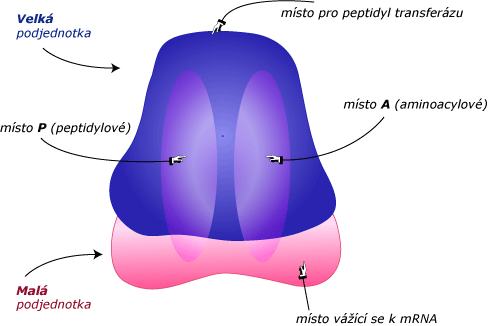
Např. malá podjednotka 30S je částí ribozomu, která se na začátku váže na mRNA. Velká 50S podjednotka se podílí na činnosti enzymu peptidyl transferázy, který katalyzuje formování peptidické vazby mezi aminokyselinami. Po spojení obou podjednotek vznikají dvě oblasti známé jako peptidylové místo (P) a aminoacylové místo (A), které zároveň váží dvě různé molekuly tRNA. Nakonec další oblasti ribozomu rozmístěných kolem obou podjednotek slouží jako místa kontaktu pro další proteiny nutné pro průběh translace.
Mechanizmus translace
Samotná translace se skládá ze tří fází:
- iniciace – začíná translace, molekula mRNA se váže na místo na ribozomu (vazebné místo pro mRNA), svou sekvencí obsahující iniciační kodon AUG,
- u prokaryot – s kodonem mRNA AUG se váží komplementárně molekuly tRNA nesoucí aminokyselinu formylmetionin (fMet) v místě P ribozomu,
- u eukaryot – se váže tRNA nesoucí aminokyselinu metionin (Met),
- tato aminokyselina se stane N koncem rostoucího polypeptidu,
- elongace – když se karboxylová skupina aminokyseliny na jedné tRNA v místě P ribozomu spojí s další aminokyselinou za vzniku peptidické vazby na druhé tRNA v místě A, ribozom se posune o 3 nukleotidy směrem k 3´konci mRNA; tento pohyb vystaví další kodon a dovolí přidání další aminokyseliny,
- tento mechanizmus zajišťuje, že směr 5´> 3´ v mRNA koresponduje se směrem N konec > C konec v rostoucím polypeptidu,
- terminace – když ribozom narazí na nesmyslný (stop) kodon, ukončuje se translace uvolnění mRNA a odpojením kompletního polypeptidu od poslední tRNA.
Posttranslační procesy mohou změnit strukturu polypeptidu
Primární struktura proteinu není pevně stanovena ještě před (probíhají kotranslační úpravy) a po skončení translace (probíhají postranslační úpravy). Probíhá zde celá řada procesů, které modifikují jeho strukturu.
Kotranslační úpravy:
- vyštěpení aminokyseliny, jako je na N konci fMet,
- přidání různých chemických látek - fosfátové skupiny, metylové skupiny, cukerné zbytky,
- tvorba disulfidických můstků.
Posttranslační úpravy (vznikají funkční proteiny):
- vyštěpení více menších peptidů z jednoho dlouhého produktu translace (polyprotein),
- tvorba kvarterní struktury a další.