Přepis genetické informace - transkripce
Tok genetické informace je vyjádřen dogmatem molekulární biologie
Přenos genetické informace je vyjádřen dogmatem molekulární biologie, které říká, že je možný přenos genetické informace z nukleové kyseliny do nukleové kyseliny, nebo z nukleové kyseliny do proteinu. Přenos informace z proteinu do nukleových kyselin, nebo z proteinu do proteinu není možný (zatím neprokázaný).
Exprese genetické informace
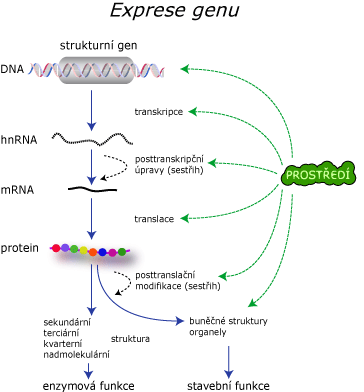
Co znamená přepis (transkripce) genetické informace?
Jedná se o přepis genetické informace z DNA do RNA. Existuje i opačný proces, zpětná transkripce, kdy se přepisuje genetická informace z RNA do DNA. Transkripcí se převádí genetická informace z formy zápisu v DNA nukleotidové sekvenci do RNA sekvence nebo naopak.
Průběh transkripce
Transkripce je první fází exprese genu. Během transkripce syntetizuje RNA polymeráza jeden komplementární řetězec primárního transkriptu z templátu DNA.
Iniciace transkripce
Enzym RNA polymeráza iniciuje transkripci, když se naváže na promotorovou sekvenci DNA a dvojitý helix se rozvine, aby se báze templátu vystavily párování s nově se tvořícím RNA řetězcem. Promotor je specializované DNA sekvence blízko začátku genu, kde začíná transkripce.
 |
 |
Typy transkripčních jednotek
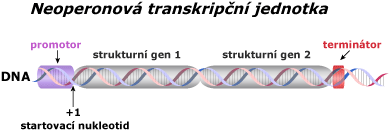
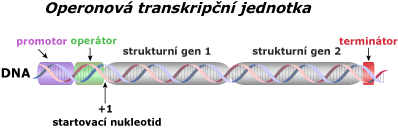
Většina bakteriálních promotorů se dělí do dvou krátkých částí, které jsou velmi konzervativní (jsou identické u většiny druhů). Tyto nukleotidy jsou místa, která rozpoznává enzym RNA polymeráza a silně se na ně váže. Většina promotorů je před prvním startovacím nukleotidem (+1). Následující příklad se vztahuje k E. Coli.
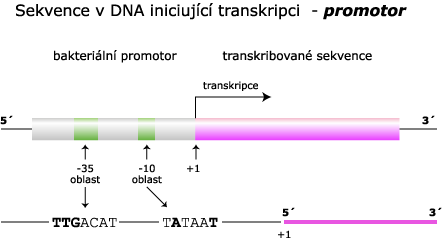
RNA polymeráza rozeznává všechny transkripční jednotky. Jedná se o holoenzym, který se skládá z několika podjednotek. Podjednotka sigma podmiňuje vazba enzymu specificky na promotor v DNA.
Po navázání na promotor rozvine RNA polymeráza ve své oblasti oba řetězce DNA a vytvoří se jednořetězcový templát DNA. Takto navázaný enzym na promotoru DNA s uvolněnými řetězci se nazývá otevřený promotorový komplex (nebo otevřený binární komplex). Enzym identifikuje templátové vlákno a vybírá dva ribonukleotidy, mezi kterými vytvoří fosfodiesterovou vazbu. Tento diribonukleotid je začátkem syntézy RNA. Sigma podjednotka se následně od DNA a enzymu odpojuje.
Elongace transkripce - konstrukce kopie RNA genu
RNA polymeráza prodlužuje mRNA ve směru 5´> 3´, když katalyzuje tvorbu fosfodiesterové vazby mezi postupně zarovnané ribonukleotidy na jejich 3´konec rostoucího RNA řetězce. Poté co se odpoutá sigma podjednotka, ztratí enzym afinitu k promotoru a získá silnou afinitu k jakékoliv sekvenci DNA. Enzym se začne pohybovat podél chromozomu, rozvíjí dvojitý helix a na uvolněném templátovém vláknu DNA syntetizuje ribonukleotidový řetězec za původním diribonukleotidem. RNA polymeráza se po templátovém vlákně DNA (negativní řetězec) pohybuje ve směru 3´> 5´. Za enzymem se obě vlákna opět spojují v dvoušroubovici. Poté co jedna RNA polymeráza opustí promotor, může na něj nasednout další enzym
Terminace transkripce - ukončení
Terminátorová sekvence v RNA zapříčiní, že RNA polymeráza se oddělí z DNA a ukončí se syntéza primárního transkriptu. Terminace se skládá ze zastavení enzymu a jeho uvolnění z DNA vlákna a RNA vlákna. Existují dva typy terminátorů:
- pravý terminátor – pro ukončení transkripce stačí jeho sekvence, které RNA polymeráza rozpozná a uvolní se,
- nepravý terminátor – pro ukončení transkripce jsou vyžadovány ještě další proteiny (ró faktor).
Terminátory často tvoří vlásenkové smyčky, v kterých se nukleotidy komplementárně spojí v páry.
Produkt transkripce je jednořetězcový primární transkript (RNA)
Podstatou primárního transkriptu jsou komplementární báze mezi místem iniciace a terminace v templátovém řetězci genu na DNA. Ribonukleotidy nesoucí tyto báze zahrnují skupiny pro startovací kodon, kodony specifikující všechny aminokyseliny v tvořeném polypeptidu při translaci a stop kodon.
U prokaryotů je primárním transkriptem přímo mRNA, která se účastní syntézy polypeptidu.
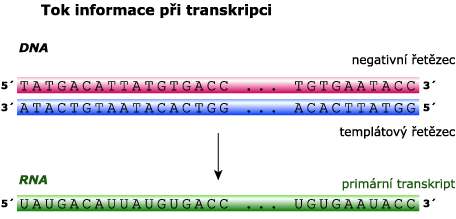
Posttranskripční úpravy primárního transkriptu se děje u eukaryotů
Protože u eukaryotů jsou také geny složené (obsahují exony i introny), musí se primární transkript (heteronukleární RNA – hnRNA; dříve se označovala jako premediátorová RNA) posttranskripčně upravit, aby vznikla zralá mRNA, která prochází z jádra do cytoplazmy, kde se účastní proteosyntézy.
Při modifikaci hnRNA se přidává metylovaná čepička na konec 5´a poly-A konec k 3´konci eukaryotické hnRNA. Modifikace neovlivňuje primární strukturu RNA a tedy genetickou informaci v ní zapsanou.
Obsahuje-li gen i introny, pak dochází ještě k sestřihu (splicing).