
![]()
![]()
|
Chromozomy obsahují mnoho genů a pokud nejsou rozděleny crossing-overem, pak alely přítomné na mnoha lokusech každého homologního chromozomu segregují jako jednotka během gametogeneze. Rekombinantní gamety jsou důsledkem crossing-overu a zvyšují genetickou variabilitu v druhu a slouží také pro konstrukci chromozomových map.
| Dosud jsme se zabývali jen volnou kombinovatelností genů, tzn. že geny jsou lokalizovány v různých nehomologních chromozomech a při procesu meiózy se náhodně kombinují. | 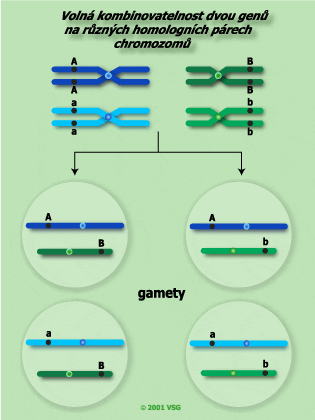 |
Počet genů jednotlivých organizmů je však vždy několikanásobně vyšší, než je počet chromozomů. Proto nemohou být všechny geny volně kombinovatelné. Ty geny, které jsou uloženy v jednom a témže chromozomu (neboli ty páry alel, které jsou neseny jedním a týmž párem homologních chromozomů) jsou vázané a jejich soubor tvoří vazbovou skupinu. Vazbou genů tedy rozumíme, že dva nebo více sledovaných genů je složkou téže vazbové skupiny. Dále se rozlišuje syntenní skupina, kdy geny na jednom chromozomu jsou tak vzdáleny, že se chovají jako volně segregující.
T. H. Morgan v roce 1926 na základě svých výzkumů definoval pravidla, která jsou označována jako Morganovy zákony:
|
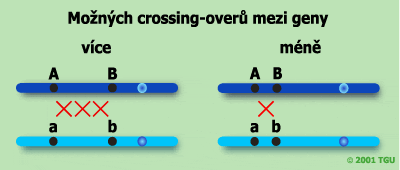
Vzdálenost mezi geny na jednom chromozomu je úměrná počtu rekombinací (crossing-overů) mezi těmito geny. Pokud bude vzdálenost větší, je větší pravděpodobnost vzniku zlomu a crossing-overu než při jejich menší vzdálenosti. Sílou vazby mezi dvěma geny rozumíme pravděpodobnost vzniku crossing-overu v oblasti, která je vymezena těmito geny. Čím jsou geny umístěny k sobě blíže, tím je vazba silnější, protože pravděpodobnost vzniku crossing-overu v dané oblasti klesá se zmenšováním její délky.
Vazbové fáze
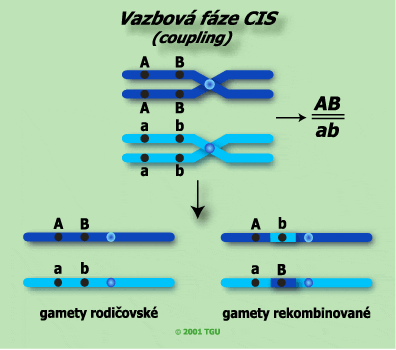
a) Vazbová fáze cis (starší označení coupling)
Při křížení rodičů o genotypech AABB a aabb, kde páry alel A/a a B/b jsou vázány, označujeme za křížení ve vazbové fázi cis. V této vazbové fázi je jeden z rodičů homozygotně dominantní v obou párech alel a druhý z rodičů v obou párech alel homozygotně recesivní:
|
|
||||||||||||||||
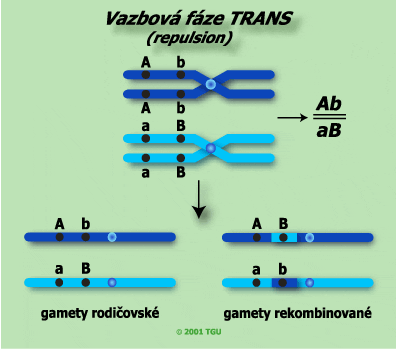
b) Vazbová fáze trans (starší označení repulsion)
Vazbou fází trans rozumíme situaci, kdy jen z rodičů je v jednom páru alel homozygotně dominantní a v druhém homozygotně recesivní (AAbb), přičemž druhý je svým genotypem vůči prvému reciproký (aaBB):
|
|
||||||||||||||||
Je třeba si uvědomit, že ne mezi všemi geny na jednom chromozomu musí dojít ke crosing-overu. Obrázek níže zaznamenává takovou situaci.
| Pokud nedojde ke crossing-overu mezi geny, tvoří se pouze dva typy různých gamet. Každá gameta získala alely, které jsou na jednom nebo na druhém homologu. Lze zde hovořit o úplné vazbě. Jejím důsledkem vznikají pouze rodičovské nerekombinované (necrosované) gamety. Oba typy gamet jsou tvořeny ve stejném poměru 1:1. | |
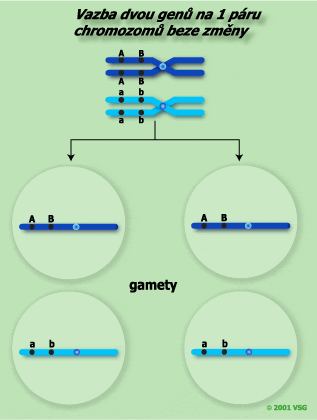
|
Zde dochází k překřížením dvou nesesterských chromatid ze čtyř ke crossing-overu. Tato výměna tvoří dvě nové kombinace alel v gametách. Ty se pak nazývají rekombinantní (crosované) gamety. |
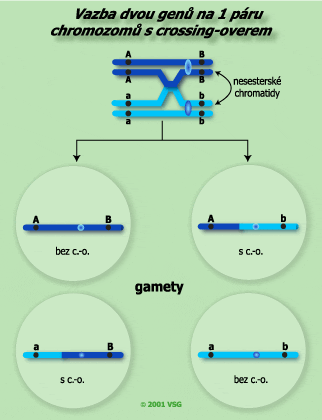
Hodnocení síly vazby genů
Pravděpodobnost,
že dva geny v dané kombinaci se nepřenesou do další generace se rovná
pravděpodobnosti výskytu crossing-overu. Tato pravděpodobnost se označuje ![]() (theta):
(theta):
|
- úplná vazba | |
| - těsná vazba | ||
| - středně těsná vazba | ||
| - volná vazba | ||
| - volná kombinovatelnost |
Vzdálenost mezi geny se vyjadřuje v centimorganech (cM), které vycházejí z rekombinačních frekvencí: 1 cM = 1 % pravděpodobnosti vzniku crossing-overů. Tato hodnota představuje mapovou vzdálenost dvou genů na chromozomu. Lze ji použít i k určení velikosti genomu nebo délky chromozomu.
Poměrnou četnost gamet rozdílných genotypů zjišťujeme hybridologickou analýzou za pomocí zpětného křížení dihybrida s recesivně homozygotním rodičem:
| Gamety rodičů | ||||
| AaBb |
AB | Ab | aB | ab |
| aabb |
ab | ab | ab | ab |
| Genotypy zygot | AaBb | Aabb | aaBb | aabb |
| Četnosti | a1 |
a2 |
a3 | a4 |
Symboly a1 až a4 označují počet zygot a tím i jedinců jednotlivých fenotypových tříd (AB, Ab, aB, ab), zároveň i počet gamet příslušných čtyř různých genotypů. Při křížení rodičů ve vazbové fázi cis mají větší četnost třídy a1 a a4, při křížení rodičů ve vazbové fázi trans třídy a2 a a3.
Vazbu můžeme charakterizovat pomocí:
Batesonovo číslo (c) – udává, kolikrát častěji vznikají gamety s nerekombinovaným uspořádáním alel oproti rekombinantním. Toto číslo však nevyjadřuje vzdálenost genů a neumožňuje jejich detekci. Proto se v současné době nepoužívá. Hodnotu c vypočítáme podle vzorce:
fáze cis |
|
fáze trans |
|
Morganovo číslo (p) – je mnohem vhodnější pro kvantifikaci síly vazby. Udává se v rekombinačních jednotkách nazvaných morgany - M (po T.H. Morganovi). Hodnota 1 centimorgan vyjadřuje, že dihybrid tvoří 1 % gamet s rekombinovanou sestavou alel; jinými slovy tzn., že v dané oblasti, vymezené sledovanými geny, je pravděpodobnost vzniku crossing-overu 1 % (0,01). Morganovo číslo lze rovněž stanovit zpětným křížením a vypočítat ve vztahu k vazbovým fázím takto:
fáze cis |
|
fáze trans |
|
Při volné kombinovatelnosti bude hodnota p rovna 50 % (0,50). Nebudou-li vznikat gamety s rekombinovanou sestavou alel, bude p rovno 0.
Vztahy mezi Batesonovým a Morganovým číslem lze zapsat:
| nebo |
Pomocí X2 testu lze zase jen zjistit, zda štěpné poměry odpovídají hypotéze volné kombinovatelnosti.
V posledních desetiletích se k vyjádření vzdálenosti genů (síly vazby) používá metoda Lod skóre. (L ~ logaritmus; od ~ odds, angl. šance, pro převahu pravděpodobnosti; skóre ~ poměru rekombinant ku nerekombinantám). Vyjadřuje logaritmus poměru pravděpodobnosti rekombinant ku nerekombinantám.
Pq - pravděpodobnost existence vazby P0,5 - pravděpodobnost neexistence vazby |
Skóre z potomstev se vypočítává v sérii
rekombinačních frakcí a výsledky jednotlivých pravděpodobností qse převedou na dekadický logaritmus, což je
vlastní Lod skóre - Z. Z různých potomstev se pak počítá jejich
součet ![]() . Je-li
. Je-li ![]()
![]() +3, pak je pravděpodobnost 1000:1 pro přítomnost vazby a naopak
+3, pak je pravděpodobnost 1000:1 pro přítomnost vazby a naopak ![]()
![]() -2 svědčí o volné kombinovatelnosti.
-2 svědčí o volné kombinovatelnosti.
Výhody Lod skóre jsou: není nutná znalost vazbové fáze; pravděpodobnost je vyjádřena v logaritmech, lze je sčítat; rekombinační frakce, v které se vypočítá maximální hodnota Z představuje vzdálenost genů.
Nejmenší lidský chromozom č. 21 je veliký 0,5 M (~50 cM) a nejdelší č. 1 má 2 M (~200 cM). Haploidní genom člověka má asi 30 M (~3000 cM) a 1 cM obsahuje kolem 1 - 2 milióny bp a představuje informační kapacitu stovek až tisíců genů. |
Porovnání vazby genů a volné kombinovatelnosti:
- pokud jsou geny na jednom chromozomu vzdáleny od sebe 50 cM, mluvíme o volné kombinovatelností; při 50 % hranici rekombinace buď nastane nebo ne, na základě hybridologické analýzy nelze tuto vazbu odlišit od volné kombinovatelnosti
- jsou-li geny umístěny na jednom chromozomu ve vzdálenosti menší než 50 cM, jsou předány potomstvu společně, dědí se pak společně, jako blok, tzn. jsou na sebe vázány. Společné dědění genů uložených na jednom chromozomu ve vzdálenosti menší jak 50 cM způsobuje v potomstvu společnou expresi vlastností determinovaných na sebe vázaných genů. Dochází tak k omezení nebo vyloučení volné kombinovatelnosti
![]()
![]()
![]()
![]()